凋落物多样性及组成对凋落物分解及土壤微生物群落的影响
——二氧化碳倍增条件下
2020-11-13王小平辛晓静曲耀冰赵念席高玉葆
王小平,杨 雪,杨 楠,辛晓静,曲耀冰,赵念席,高玉葆
南开大学生命科学学院,天津 300071
自工业革命以来,化石燃料的大量燃烧和森林的过度采伐、草地等植被的破坏导致大气CO2浓度以前所未有的速度增加[1]。在过去几十年中,CO2浓度变化如何影响植物的生长和竞争[2]、群落的组成和结构[3]等逐渐受到研究者的关注,如Zuo等[4]在美国明尼苏达州中东部的雪松溪生态系统科学保护区进行了为期4年的对比实验发现,相对于较低CO2浓度处理,高CO2浓度条件下培养所得植物的凋落物N含量更低;Cotrufo等[5]在英国坎布里亚州米托普伍德混合落叶灌木林中对凋落物分解进行了为期1年的研究发现,与当前大气CO2浓度(400 μmol/mol)相比,高CO2浓度条件下凋落物C/N值更高,凋落物分解速率更低;并得出凋落物初始C/N与凋落物分解呈负相关关系,其中,凋落物初始C/N可作为预测凋落物分解速率的重要指标[6- 7]。此外,也有一些研究者比较关注CO2浓度升高如何影响土壤微生物群落,如Kandeler等[8]在美国科罗拉多州草原经过5年的研究发现,CO2浓度升高可提高土壤真菌的PLFAs含量;但Hagedorn等[9]在斯蒂尔伯格的高寒森林中进行为期9年的研究发现,CO2浓度变化对土壤微生物含量和群落组成影响很小。
受全球变化的影响,生物多样性的丧失日益严重,势必会对物质循环这一生态系统功能产生显著影响。因此,凋落物物种多样性及组成如何影响凋落物分解及土壤微生物群落已受到研究者的广泛关注,并取得了系列有科学价值的研究成果[4,10- 13]。然而,这些研究均在当前大气CO2浓度下开展,并未考虑环境CO2浓度变化对物种多样性和凋落物分解之间关系的影响。有研究预测,受人类强烈活动的影响,到2100年大气CO2浓度将超过800 μmol/mol[14],因此,在高于当前大气CO2浓度条件下,探索不同物种及不同物种组合对凋落物分解特性和土壤微生物群落的影响,一方面可为预测全球变化导致的生态后果进行准确评估。另一方面与当前大气CO2浓度下所得结果进行对比,还可为环境CO2浓度变化对生态系统物质循环的影响及其作用机制提供数据支持。
草地生态系统是我国陆地生态系统最重要的类型之一,其面积占陆地面积的40%[15]。受全球变化的影响,草原群落退化严重,表现为群落物种多样性降低、建群种更替等,进而影响该区域生态系统功能。如王小平等[16]以呼伦贝尔草原区一处退化恢复样地中的4种主要植物为研究对象,在当前大气CO2浓度条件下,发现凋落物的初始性质(初始C、N、木质素含量及初始C/N)会对凋落物质量、C、N残余率及残余C/N的一种或几种具有显著的直接影响,还可通过土壤真菌PLFAs含量显著影响凋落物质量残余率。为了深入探讨全球变化对草原凋落物分解这一生态系统功能的影响,本研究在两倍于当前大气CO2浓度(800 μmol/mol)条件下,来研究凋落物物种多样性及其组成对凋落物分解及土壤微生物群落的影响,并重点关注:凋落物多样性及组成与凋落物分解以及土壤微生物群落的关系是怎样的?这种关系与当前大气CO2浓度条件下所得结果有哪些异同点?
1 材料与方法
1.1 实验材料
2014年8月初,在内蒙古自治区呼伦贝尔市鄂温克族自治旗北辉野外科学观测研究站的一个围封样地内(48.15°—49.00°N,118.80°—119.75°E)采样。该样地原生植被为贝加尔针茅(Stipabaicalensis)-羊草(Leymuschinensis)草原,由于过度放牧,草原退化严重(建群种更替),于2008年开始围封禁牧。基于群落调查的物种重要值排序结果,收集重要值高的羊草、茵陈蒿(Artemisiacapillaris)、麻花头(Serratulacentauroides)和二裂委陵菜(Potentillabifurca)4种植物的立枯凋落物,风干至恒重;并用土钻收集地表(0—10 cm)新鲜土壤,用于室内凋落物分解。该区域位于内蒙古草原区的东北部,温带大陆性气候,属于半干旱区,年降水量250—350 mm,年气候特征表现为冬季寒冷干燥,夏季炎热多雨,土壤类型为暗栗钙土。
本实验所用4种凋落物的初始性质同王小平等[16]一致。羊草、茵陈蒿、麻花头和二裂委陵菜凋落物的初始全C含量(C0)依次为39.63%、38.05%、37.30%和37.93%;初始全N含量(N0)依次为1.50%、0.95%、0.75%、1.01%;初始木质素含量依次为16%、19%、31%和32%;羊草凋落物具有最高的初始全N含量,最低的初始木质素含量和初始C/N。
1.2 实验设计
本实验采用嵌套实验设计,将凋落物组成嵌套在凋落物多样性内。其中凋落物多样性,包括1、2、4物种三个水平;物种组合共11种,详见表1。每种凋落物组合共1.200 g(M0),装入网孔面积为1 mm2凋落物袋中,多物种组合中各个物种组分凋落物质量相等,每种组合3个重复,共33袋。

表1 凋落物多样性及凋落物组成方式(不同处理以分号隔开)
凋落物分解实验于智能人工气候箱中进行:将凋落物袋置于培养皿(1袋/皿)正上方,凋落物袋下方为50 g原生生境土壤。人工气候箱设置12 h光照、光强为60 μmol m-2s-1、湿度为50%、温度为25℃和CO2浓度设置为800 μmol/mol。实验期间,每3d随机交换培养皿位置以避免位置效应。60 d后,收获每个凋落物袋中的凋落物,测定剩余凋落物质量(MT)、全C(Ct)、全N(Nt)含量用于残余率分析,同时采集每个培养皿中的土壤用于土壤微生物组成及结构分析。
1.3 实验方法
利用Vario MICRO cube元素分析仪测定凋落物初始及实验结束时全C、全N含量,并计算对应的C/N;参考Antonietta等[17]方法测定凋落物的初始木质素含量。
采用磷脂脂肪酸(Phospholipid Fatty Acid,PLFA)温和甲酯化法[18]和气质联用色谱分析仪(Agilent 7890GC,5975MSD)进行土壤微生物PLFA生物标记分离、鉴定及含量计算[19],并依据相关文献对所得PLFA表征分类[20- 22],包括细菌(区分革兰氏阳性菌G+、革兰氏阴性菌G-和一般细菌)、真菌以及计算的真菌/细菌(F/B)。
1.4 数据分析
依据Mt=MT/M0×100%,MC=Mt×Ct/C0和MN=Mt×Nt/N0分别计算凋落物质量、C、N残余率(%)。
利用线性混合模型(Linear Mixed Model)的嵌套单因素方差分析来研究凋落物多样性及组成(其中,凋落物组成嵌套在凋落物多样性内)对凋落物质量、C、N残余率及残余C/N和土壤微生物各类群PLFAs含量影响的显著性(SPSS 21.0);借助冗余分析(Redundancy Analysis,RDA)来研究凋落物多样性及组成与凋落物质量、C、N残余率、残余C/N及土壤G+、G-及真菌PLFAs含量之间的关系(Canoco for Windows 4.5);利用“sem”、“dplyr”及“DiagrammeR”程序包构建结构方程模型(Structural Equation Model,SEM)来分析预测变量,包括初始C、N、木质素含量及初始C/N,G+、G-、真菌PLFAs含量及F/B,与凋落物分解相关参数(凋落物质量、C、N残余率及残余C/N)之间的关系(R 3.5.1)。
2 结果与分析
2.1 凋落物分解
凋落物多样性对凋落物质量、C、N残余率及残余C/N均有显著影响(P<0.05)(图1)。其中,4物种凋落物多样性处理下,凋落物质量、C残余率均显著低于其他两种处理所得结果,凋落物N残余率与其他两种处理所得结果无显著差异,而残余C/N仅显著低于单物种多样性处理;相对于单物种凋落物多样性处理,2物种凋落物多样性处理表现出显著高的N残余率和显著低的残余C/N。
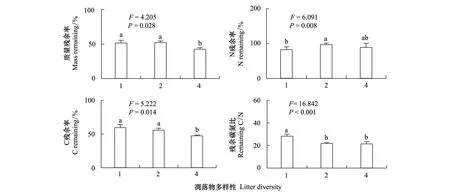
图1 凋落物多样性对凋落物质量、C、N残余率及残余C/N的影响Fig.1 The effects of litter diversity on litter mass,C,N remaining rate and remaining C/N图中相同的英文字母表示处理间差异不显著
凋落物组成对凋落物质量、C和N残余率均具有显著影响(P<0.05)(图2)。单物种凋落物多样性组合中,茵陈蒿(A)的凋落物质量、C和N残余率均显著(P<0.05)低于其余3种植物;2物种凋落物多样性组合中,麻花头+二裂委陵菜(S+P)的质量残余率显著(P<0.05)高于除茵陈蒿+麻花头(A+S)外的组合,而C残余率显著高于其他五种组合;4物种凋落物多样性组合(L+A+S+P),其质量残余率显著低于除茵陈蒿(A)、羊草+茵陈蒿(L+A)及茵陈蒿+二裂委陵菜(A+P)外的其他组合;其C残余率显著低于除茵陈蒿(A)、羊草+麻花头(L+S)、羊草+茵陈蒿(L+A)及茵陈蒿+二裂委陵菜(A+P)外的其他组合;其N残余率与麻花头(S)、茵陈蒿(A)、麻花头+二裂委陵菜(S+P)及茵陈蒿+麻花头(A+S)组合间均存在显著差异。
凋落物组成对凋落物残余C/N具有显著影响(P<0.05)。单物种凋落物多样性组合中,羊草(L)的残余C/N显著(P<0.05)低于其他3种植物;2物种凋落物多样性组合中,麻花头+二裂委陵菜(S+P)的残余C/N显著(P<0.05)高于除茵陈蒿+二裂委陵菜(A+P)外的其他组合;4物种凋落物多样性组合(L+A+S+P)所得残余C/N显著(P<0.05)低于单物种麻花头(S)、二裂委陵菜(P)及茵陈蒿(A)组合处理。
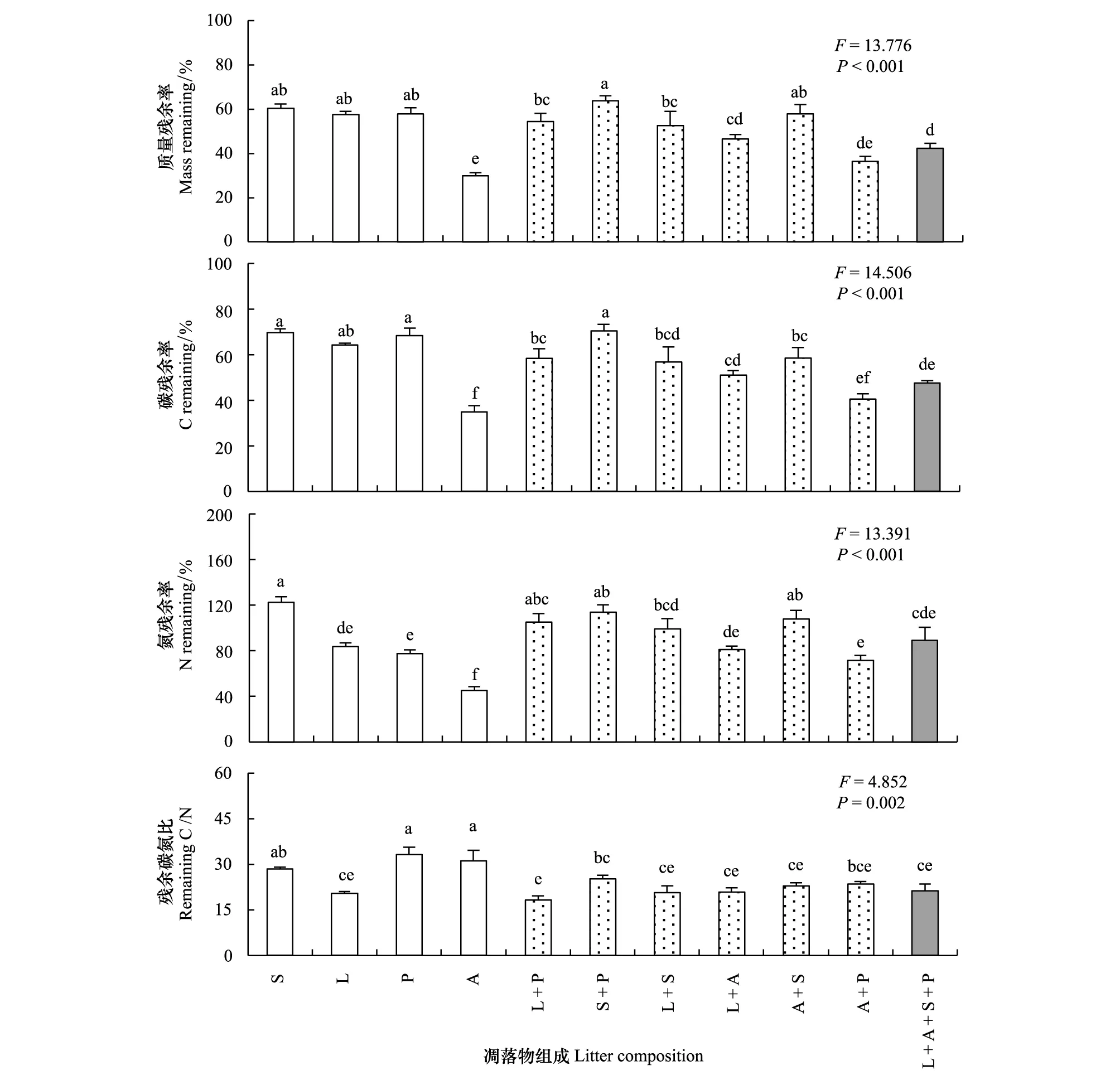
图2 凋落物组成对凋落物质量、C、N残余率及残余C/N的影响Fig.2 The effects of litter composition on litter mass remaining,C remaining,N remaining and remaining C/NS: 麻花头 S. centauroides;L:羊草 L. chinensis;P:二裂委陵菜 P. bifurca;A: 茵陈蒿 A. capillaris
2.2 土壤微生物群落组成和结构
凋落物多样性对所观测的土壤微生物群落组成和结构指标均无显著(P>0.05)影响(表2);凋落物组成对G+PLFAs含量具有显著(P<0.05)影响(表2),具体表现为:单物种凋落物多样性组合所得土壤中,麻花头(S)凋落物下土壤的G+PLFAs含量显著(P<0.05)高于羊草(L)及二裂委陵菜(P)下土壤所得结果;2物种凋落物多样性组合所得土壤中,羊草+二裂委陵菜(L+P)下土壤的G+PLFAs含量显著高于麻花头+二裂委陵菜(S+P)、茵陈蒿+麻花头(A+S)以及茵陈蒿+二裂委陵菜(A+P)凋落物下土壤所得结果;4物种凋落物多样性组合(L+A+S+P)所得土壤的G+PLFAs含量显著(P<0.05)低于麻花头(S)、茵陈蒿(A)、羊草+二裂委陵菜(L+P)以及羊草+麻花头(L+S)下土壤所得结果(图3)。
2.3 冗余分析
冗余分析结果表明:前两轴共解释了50%的变异,且排序轴对应的环境变量对响应变量的解释贡献达显著水平(F=3.44,P=0.014),对总变异贡献的大小依次为茵陈蒿(28%)>麻花头(21%)>凋落物多样性(10%)>羊草(8%)>二裂委陵菜(7%)。其中,凋落物质量、C、N残余率与含凋落物茵陈蒿的处理呈显著负相关;凋落物质量、C、N残余率以及土壤微生物G+、G-和真菌(PLFAs)含量均与含凋落物麻花头及二裂委陵菜的处理呈显著正相关;真菌、G+、G-、PLFAs及残余C/N与凋落物多样性和含凋落物羊草的处理呈显著负相关(图4)。
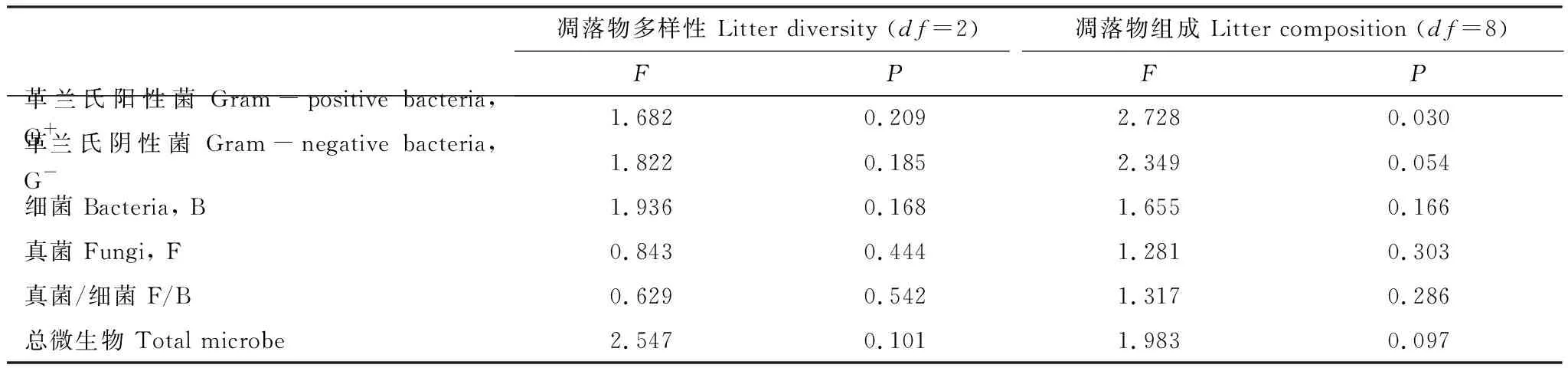
表2 凋落物多样性及组成对土壤微生物群落组成和结构影响的嵌套式分析

图3 凋落物组成对土壤G+PLFAs含量的影响Fig.3 The effects of litter composition on the soil G+ PLFAs contentS: 麻花头 S. centauroides;L:羊草 L. chinensis;P:二裂委陵菜 P. bifurca;A: 茵陈蒿 A. capillaris
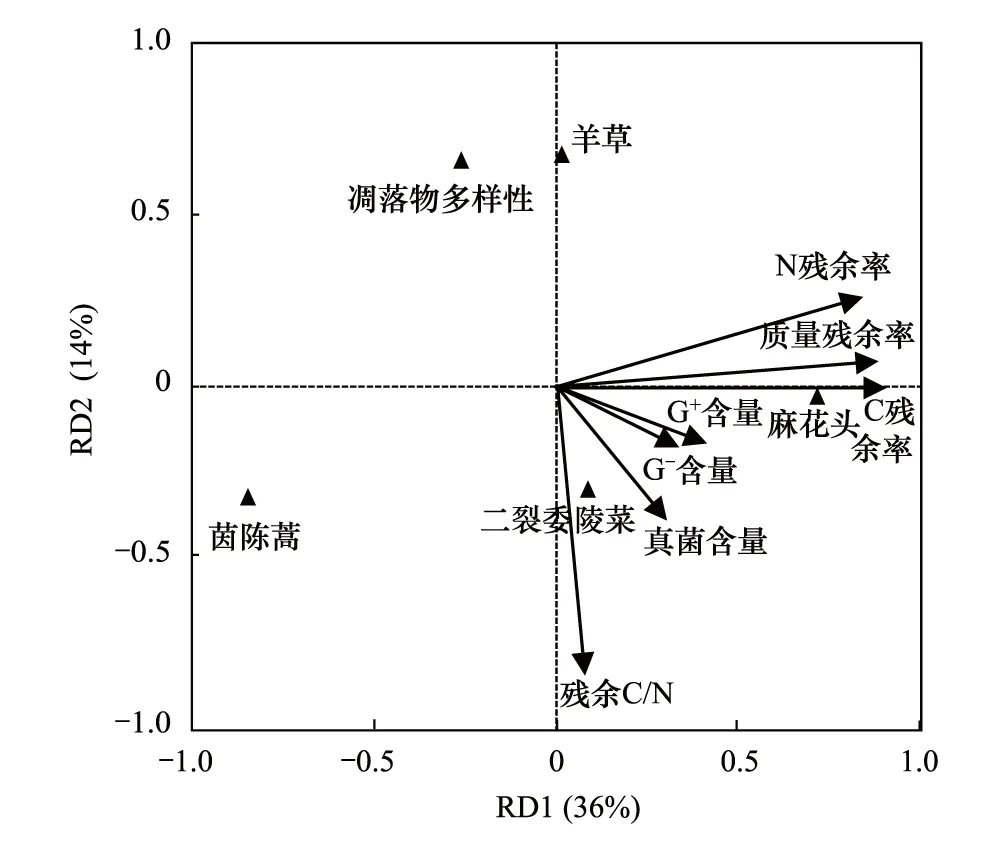
图4 土壤微生物群落和凋落物分解特性与凋落物多样性及组成相关关系的冗余分析Fig.4 The relationships between soil microbial community and characteristics of litter decomposition and litter diversity and composition by redundancy analysis
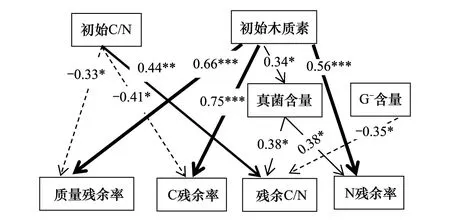
图5 凋落物初始化学性质、土壤微生物群落与凋落物分解特性的结构方程模型Fig.5 The structural equation model of initial litter chemical properties,soil microbial community and litter decomposition characteristics 图中仅显示P<0.05的相关关系,实线、虚线箭头分别表示正、负相关关系,*:P<0.05,**:P<0.01,***:P<0.001
2.4 结构方程模型
经显著性检验,所得结构方程模型结果表明:凋落物初始C/N对凋落物质量、C残余率及残余C/N具有显著直接影响。然而,凋落物初始木质素含量对凋落物质量、C及N残余率具有显著直接影响,也可通过对真菌PLFAs含量的影响来间接对凋落物N残余率及残余C/N产生显著影响。G-PLFAs含量对残余C/N具有显著直接影响(图5)。
3 讨论
干旱-半干旱区草原地上部分,除牲畜采食外,绝大部分以凋落物的形式参与生态系统物质循环。因此,草原群落的种类组成和结构对凋落物的质量具有决定性的作用,对维护草原土壤质量和发挥草原生态系统功能方面具有重要作用。草原退化导致建群种替代,即使经过多年恢复,与原生群落的物种组成间仍有差异。本研究区域为经过6年围封的恢复群落,群落中优势度较大的以杂类草麻花头、二裂委陵菜和茵陈蒿为主,与原生植被以贝加尔针茅、羊草为主导的群落物种组成有较大差异。这些杂类草初始凋落物具有高的初始C/N和木质素含量,且这些因素主导并减缓凋落物分解进程,如王小平等在当前大气CO2浓度条件下研究指出,该区域生态系统物质循环较原生群落慢[16]。而本研究发现,在两倍于当前大气CO2浓度条件下,凋落物的分解速率较王小平等[16]于当前大气CO2浓度条件下所得结果更慢,这与高CO2浓度下凋落物质量、C残余率高,进而导致残余C/N高有关。可能的原因为(1)凋落物残余C/N增加造成微生物生物量受限制,凋落物分解速率将会变低[23]。但这种影响具有物种组成的特异性,如茵陈蒿凋落物,其土壤中G+含量较丰富(图3),但凋落物残余率则较低(图2)。(2)CO2浓度升高除了影响凋落物化学组成及土壤微生物外,还会引起土壤pH、温度及土壤动物等因素的变化[24-25],从而直接或间接地降低凋落物分解速率,如Cotrufo等[5]发现高CO2浓度环境中土壤动物种群数量相对较少。但也有与此结果不同的报道,如Conteaux等[26]研究发现高CO2浓度环境下C/N升高的凋落物,仅在初期分解减慢,随后加快;Williams等[27]在研究曼哈顿的高草原凋落物分解时发现CO2浓度升高,土壤微生物分解活动增加,促进了凋落物分解。这些结果表明大气CO2浓度升高对生物、以及凋落物分解的影响是非常复杂的,观测区域及凋落物分解时长均会影响观测结果。
本研究发现,在两倍于当前大气CO2浓度条件下,凋落物组成对凋落物分解各参数及土壤微生物群落指标的影响高于凋落物多样性(图4),结构方程模型(图5)支持凋落物初始木质素含量及初始C/N对凋落物分解参数有显著直接的影响,且凋落物初始木质素含量起主导作用,为群落物种替代对生态系统物质循环的显著影响提供了直接的证据。此外,研究还发现G-含量显著影响凋落物残余C/N,原因可能为CO2浓度变化影响凋落物分解过程中的组分变化[28],因而满足了G-的需求,G-倾向于高质量(如低C/N,低木质素含量)的凋落物环境[29],凋落物羊草(低C/N,低木质素含量)可能为其提供了条件。这一结果与当前大气CO2浓度条件所得结果不一致[16],但并不影响凋落物初始木质素含量对凋落物分解的主导地位。刘书林等[30]通过全自动微气候控制的“人工模拟气候实验系统”对川西亚高山针叶林土壤进行模拟CO2倍增对土壤中微生物群落结构的影响,得出高CO2浓度对G+有显著影响;Liu等[31]以稻田生态系统为研究对象,发现CO2浓度升高显著增加了水稻土壤中真菌的丰度。
本研究结果为全球气候变化大背景下,退化恢复草原生态系统的物质循环特征提供了基础数据,也可为相似生态系统的生态系统功能的合理评估提供数据参考。
