熊果酸靶向自噬促进小鼠C2C12成肌细胞增殖①
2020-11-10张晓荣李梦俊田嵩浩高艳萍
张晓荣 王 婧 李梦俊 田嵩浩 高艳萍
(山西医科大学汾阳学院,汾阳 032200)
骨骼肌卫星细胞是位于骨骼肌基底膜和肌膜之间的成体祖细胞,可表达配对盒蛋白3(paired box 3,PAX3)、配对盒蛋白7(paired box 7,PAX7)等。当骨骼肌受到刺激时,PAX7阳性的卫星细胞被激活,生成大量的成肌细胞,为骨骼肌的维持、再生、修复提供肌核与肌纤维。小鼠C2C12成肌细胞是来源于骨骼肌卫星细胞的肌源细胞,广泛用于肌肉再生的体外模型研究。大量研究表明,癌症、恶病质、糖尿病、败血症、衰老等可导致骨骼肌干细胞衰老、肌源细胞干性和功能丧失、肌纤维丢失,骨骼肌大量丢失可引起肌肉萎缩、失能,甚至死亡[1]。因此,对肌源细胞的正向调控是防治肌肉萎缩相关疾病的基础。而目前临床上缺乏有效预防和治疗骨骼肌衰老萎缩的药物。熊果酸是一种五环三萜类天然化合物,大量研究发现其对肥胖、糖尿病、抗炎症、抗氧化反应及乳腺癌、肺癌等肿瘤都有作用[2-5]。有研究表明在饥饿诱导的肌肉衰老模型中,熊果酸可以抑制肌肉衰老相关基因的表达,抑制骨骼肌衰老[6]。此外,熊果酸可以促进肌肉衰老大鼠的腓肠肌量增多[7]。熊果酸可以刺激胰岛素样生长因子(insulin like growth factor-Ⅰ,IGF-Ⅰ)的表达,增强蛋白激酶B(protein kinase B,PKB/AKT)、哺乳动物雷帕霉素受体(mammalian target of rapamycin,mTOR)、AMP依赖的蛋白激酶(AMP-activated protein kinase,AMPK)等表达促进肌肉肥大[8]。熊果酸对肌肉调控作用的机制目前尚不清楚,本实验通过研究熊果酸对C2C12成肌细胞的调控作用及机制,为骨骼肌萎缩的治疗及预防提供新的策略和参考。
1 材料与方法
1.1材料 C2C12细胞株由天津医科大学牛凯军教授惠赠;熊果酸购自南京道斯夫生物公司;3-Methyladenine(3-MA,HY-19312)购自MCE公司;细胞培养基、CCK8试剂盒、二抗、DAPI染色液、荧光二抗购自博士德公司;EdU-488细胞增殖试剂盒购自碧云天公司;细胞周期检测试剂盒购自索莱宝公司;PAX7、P62、LC3B、P16单克隆抗体均购自Abcam公司。
1.2方法
1.2.1细胞培养及药物处理 C2C12细胞于含10%胎牛血清和1%青-链霉素的DMEM培养基、37℃、5% CO2恒温培养箱培养。不同浓度(0.1、0.5、1.5、10 μg/ml)熊果酸处理后利用CCK8检测细胞增殖情况。研究熊果酸(0.5、1 μg/ml)对细胞自噬水平的影响,后续实验选用1 μg/ml熊果酸处理细胞。
1.2.2CCK8细胞增殖实验 C2C12细胞以初始密度1×104个/孔接种到96孔板中,边缘孔用PBS覆盖。24 h后用不同浓度熊果酸(0.1、0.5、1、5、10 μg/ml)培养细胞24 h,每孔加入10 μl CCK8溶液后继续孵育1 h。酶标仪检测450 nm处的吸光度(OD)值。
1.2.3Western blot 熊果酸处理过的细胞用PBS清洗后,加入含蛋白酶抑制剂的细胞裂解液提取总蛋白,BCA试剂盒对蛋白定量,按照40 μg/孔的蛋白量进行12% SDS-PAGE凝胶电泳,蛋白分离后转至NC膜,5%脱脂牛奶室温封闭2 h,加入相应的一抗在4℃过夜,使用TBST洗膜后,加入二抗室温孵育2 h,TBST洗膜,最后加入发光液,于凝胶成像仪曝光拍照并统计灰度值,计算PAX7、P62、LC3B、P16蛋白相对表达量。
1.2.4EdU-488检测细胞增殖 6孔板中熊果酸和3-MA处理完成后,加入EdU工作液继续孵育 2 h,去除培养液后,每孔加入1 ml固定液,室温固定15 min。去除固定液,用3% BSA的PBS洗涤 3次。弃洗涤液,0.3%Triton X-100室温孵育15 min,洗涤液洗涤3次。弃洗涤液,每孔加入500 μl Click反应液室温避光孵育30 min,洗涤液洗涤3次。弃洗涤液,每孔加入1 ml Hoechst室温避光孵育10 min,洗涤液洗涤3次后用荧光显微镜观察。
1.2.5流式细胞术检测细胞周期 细胞经熊果酸和3-MA处理后,收集1×106个细胞并用预冷70%乙醇4℃过夜固定。第2天用PBS洗去固定液,4℃以1 000 r/min离心10 min,沉淀中加入100 μl RNase A溶液重悬,37℃水浴30 min。避光条件下,每管加入400 μl PI,4℃孵育30 min,上流式细胞仪检测细胞周期。
1.2.6免疫荧光 细胞经熊果酸和3-MA处理后,PBS洗涤3次,每次5 min。用预冷的4%多聚甲醛固定20 min,PBS洗涤3次,每次5 min。用0.5%Triton X-100室温通透20 min,1% BSA室温封闭1 h后,加入一抗4℃孵育过夜。第2天用荧光二抗室温避光孵育1 h,PBS洗涤3次后,DAPI室温孵育10 min,PBS洗涤3次后荧光显微镜观察P62表达。

2 结果
2.1熊果酸对C2C12细胞生长增殖能力的影响 采用CCK8方法,检测不同浓度(0.1、0.5、1、5、10 μg/ml)熊果酸对C2C12细胞增殖的影响。当熊果酸浓度<1 μg/ml时促进C2C12细胞增殖,浓度>5 μg/ml时,熊果酸对细胞生长增殖的促进作用消失,并在10 μg/ml高浓度组表现出细胞毒性作用。与Control组相比,10 μg/ml熊果酸组C2C12细胞贴壁数目减少,细胞密度明显降低,背景中可见大量细胞碎片,细胞萎缩并逐渐变圆,胞质减少,见图1。

图1 不同浓度熊果酸对C2C12细胞增殖能力的影响Fig.1 Effects of ursolic acid with different concentrations on C2C12 cells proliferationNote:*.P<0.05 vs Control.
2.2Western blot检测熊果酸对C2C12细胞中PAX7蛋白表达的影响 Western blot检测0.5、1 μg/ml熊果酸干预的C2C12细胞中PAX7蛋白的表达情况。如图2所示,与Control组相比,熊果酸(0.5、1 μg/ml)组PAX7蛋白表达量明显增加[81.20±6.39,86.57±6.84 vs 47.43±3.31],差异有统计学意义(P<0.05)。
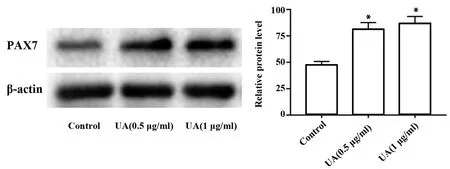
图2 Western blot检测PAX7蛋白表达Fig.2 Expression of PAX7 protein was tested by Western blotNote:*.P<0.05 vs Control.
2.3熊果酸对自噬相关蛋白表达的影响 通过Western blot实验检测熊果酸对C2C12细胞自噬相关蛋白表达的影响。如图3所示,与Control组相比,熊果酸(0.5、1 μg/ml)组P62蛋白表达量明显下降[83.56±11.72,76.48±7.83 vs 89.83±10.04],差异有统计学意义(P<0.05);LC3Ⅱ/LC3Ⅰ蛋白表达显著增加[79.56±1.45,117.06±2.49 vs 53.81±0.83],差异有统计学意义(P<0.05)。
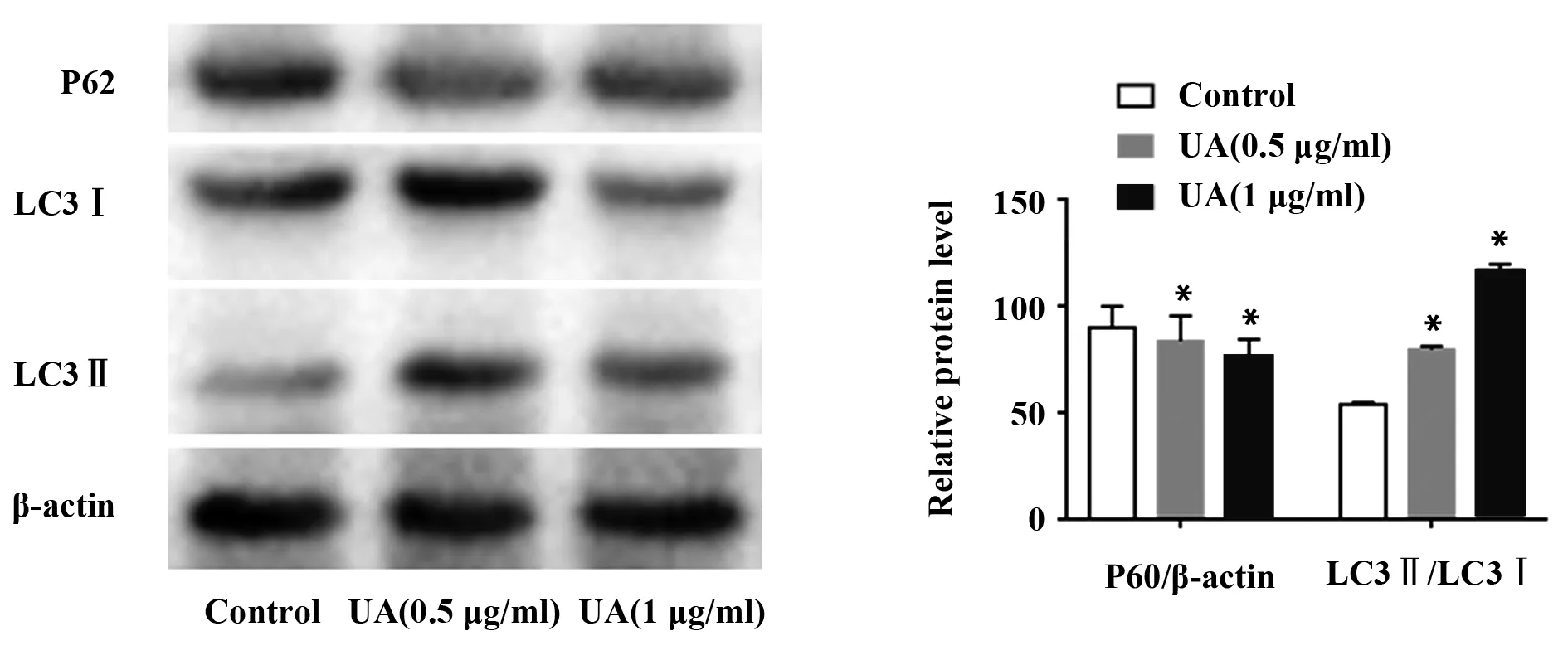
图3 Western blot检测细胞自噬相关蛋白表达Fig.3 Expressions of autophagy-related proteins were tested by Western blotNote:*.P<0.05 vs Control.
2.43-MA抑制自噬水平 使用1 mmol/L 3-MA提前1 h预处理细胞,采用Western blot和免疫荧光实验检测自噬水平变化。如图4A、表1所示,与Control组相比,3-MA组P62蛋白表达上升,LC3Ⅱ/LC3Ⅰ表达下降。与UA组相比,UA(1 μg/ml)+3-MA组P62蛋白表达上升,LC3Ⅱ/LC3Ⅰ表达上升,表明自噬流被阻断。与3-MA组相比,UA(1 μg/ml)+3-MA组P62蛋白表达下降,LC3Ⅱ/LC3Ⅰ表达上升。如图4B中免疫荧光结果所示,与Control组相比,3-MA组P62在胞质中颗粒聚集。与3-MA组相比,UA(1 μg/ml)+3-MA组P62荧光聚集减少,荧光强度降低。
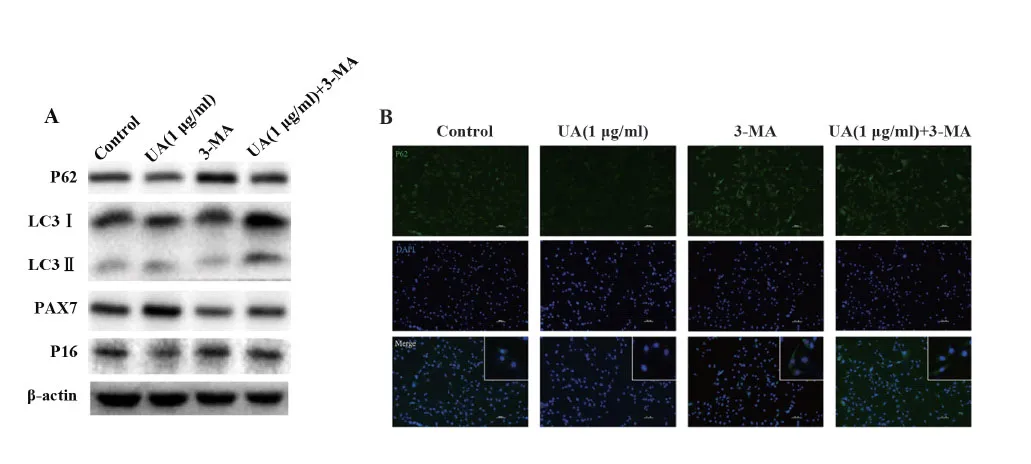
图4 Western blot和免疫荧光检测蛋白表达Fig.4 Expression of proteins were tested by Western blot and immunofluorescence assay

表1 各组细胞相关蛋白表达Tab.1 Expressions of cell-related proteins in different
2.5熊果酸通过自噬促进C2C12细胞增殖 通过EdU-488检测自噬抑制后,熊果酸对C2C12细胞增殖的影响。图5结果显示,与Control组相比,UA(1 μg/ml)组绿色荧光细胞数明显增加[50.87±1.44 vs 36.75±4.31]。与UA(1 μg/ml)组相比,3-MA组绿色荧光细胞数减少[33.85±1.37 vs 50.87±1.44;P<0.05],UA (1 μg/ml)+3-MA组绿色荧光细胞数减少[37.32±3.99 vs 50.87±1.44;P<0.05],而3-MA组与UA (1 μg/ml)+3-MA组绿色荧光细胞数相比差异无统计学意义(P>0.05)。
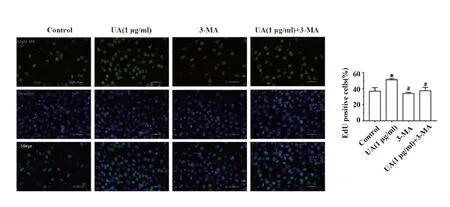
图5 EdU-488检测细胞增殖改变Fig.5 Cell proliferation was detected by EdU-488Note: *.P<0.05 vs Control;#.P<0.05 vs UA(1 μg/ml).
2.6熊果酸通过自噬促进PAX7蛋白表达 Western blot检测熊果酸和3-MA干预的C2C12细胞中PAX7蛋白的表达情况。如图4A、表1所示,与UA(1 μg/ml)组相比,UA(1 μg/ml)+3-MA组PAX7蛋白表达量明显减少,但其表达量较3-MA组增加,差异有统计学意义。
2.7熊果酸通过自噬影响细胞周期 熊果酸和3-MA干预C2C12细胞后,Western blot检测P16蛋白的表达情况,流式细胞仪检测细胞周期变化。如图4A、表1所示,UA(1 μg/ml)组P16蛋白表达量较Control组减少;UA(1 μg/ml)+3-MA组的P16蛋白表达量较Control组和UA(1 μg/ml)组显著增加。如图6、表2所示,UA(1 μg/ml)组G1/G0期细胞较Control组减少,UA(1 μg/ml)+3-MA组G1/G0期细胞较UA(1 μg/ml)组增加。
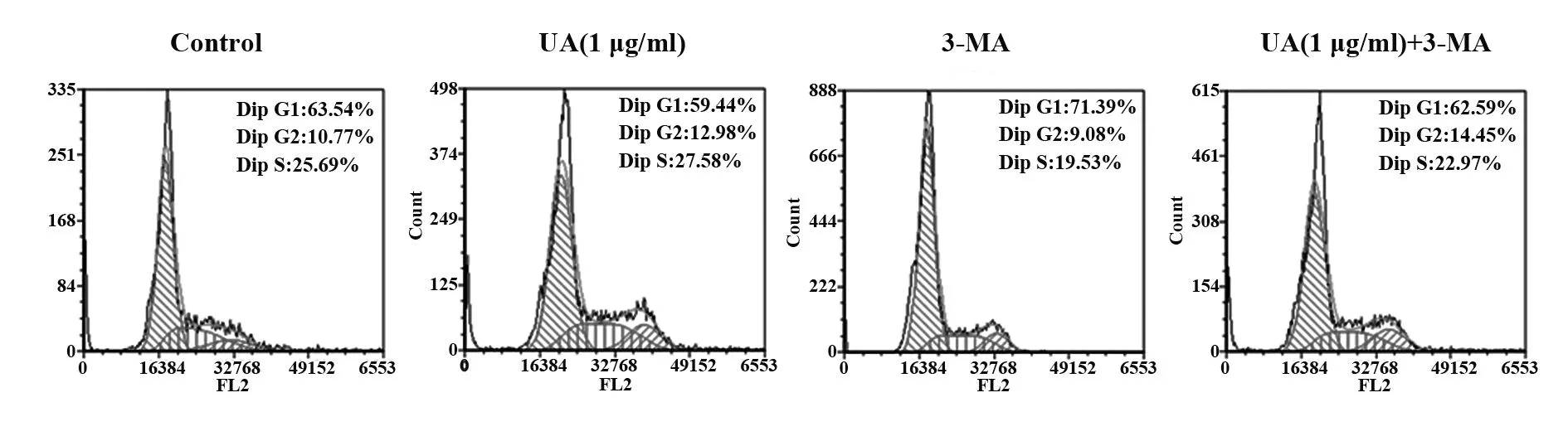
图6 流式细胞仪检测细胞周期时相分布Fig.6 Cell cycle phase was examined by flow cytometry
表2 流式细胞术检测各组细胞周期分布Tab.2 Cell cycle distribution of each group was detected by flow
3 讨论
多种慢性疾病、消耗性疾病以及衰老等容易引起骨骼肌萎缩,导致骨骼肌功能丧失甚至死亡。骨骼肌卫星细胞作为骨骼肌的干细胞,对骨骼肌的发育、维持和修复至关重要,近年来有大量研究通过异体移植肌肉卫星细胞以修复骨骼肌的损伤[9]。C2C12成肌细胞是来源于卫星细胞的肌源细胞,可以增殖、分化为肌管和肌纤维,是体外研究肌肉再生的最佳模型。
熊果酸是广泛存在于苹果皮中的化合物,具有促进癌细胞凋亡的作用。有研究发现熊果酸对热应激损伤的心肌细胞具有抗凋亡和抗氧化作用[5]。此外,熊果酸可以抑制骨骼肌衰老的相关基因的表达,促进肌肉肥大,但其调控机制目前尚不明确。骨骼肌卫星细胞是维持和修复骨骼肌的关键细胞,本研究通过不同浓度熊果酸干预小鼠C2C12成肌细胞,发现浓度小于1 μg/ml的熊果酸具有促进细胞增殖的作用。PAX7是C2C12成肌细胞增殖所必需的蛋白,在骨骼肌干细胞功能的维持和骨骼肌的再生中发挥重要作用[10]。本研究发现熊果酸(0.5、1 μg/ml)可以促进PAX7蛋白表达,增强C2C12细胞增殖能力。高浓度熊果酸(10 μg/ml)对C2C12成肌细胞有细胞毒性,可能与凋亡、焦亡或巨胞饮等非凋亡性细胞死亡有关,其毒性机理有待进一步研究,而在开发熊果酸相关药物时需注意高剂量药物毒性的影响[11]。
自噬是生物体内重要的生物降解程序,可以将受损的细胞器、蛋白、脂质降解以维持细胞稳态。自噬在维持肌肉和肌纤维的完整性中发挥重要的作用,基础自噬有利于维持骨骼肌干细胞的干性、抑制衰老。自噬功能受损可能是导致肌纤维退化、肌肉萎缩的原因之一,而在肌少症中增强自噬可以维持肌肉卫星细胞的干性,促进肌肉再生[12,13]。有研究表明熊果酸通过增强自噬活性以调控TLR4/MyD88信号通路,抑制炎症[14]。熊果酸还通过激活自噬抑制人肺癌细胞A549[15]。P62是细胞自噬底物蛋白,可以靶向其他蛋白进行选择性的自噬;微管相关蛋白1轻链3(LC3)参与自噬体膜延伸以及自噬溶酶体降解过程。本研究发现熊果酸(0.5、1 μg/ml)促进P62蛋白的降解,抑制P62蛋白的表达,促进LC3Ⅱ蛋白表达,表明熊果酸可以增强细胞自噬活性。
为研究熊果酸对C2C12成肌细胞增殖的促进作用是否靶向自噬,本研究通过自噬抑制剂3-MA阻断自噬,P62、LC3蛋白结果表明3-MA预处理抑制自噬活性,3-MA预处理的熊果酸组LC3Ⅱ/LC3Ⅰ和P62表达增加,胞质中P62荧光颗粒聚集,自噬溶酶体降解过程被抑制,自噬流受阻。3-MA抑制自噬水平后,通过EdU实验发现,EdU掺入显著减少,熊果酸对增殖的促进作用被抑制;此外,细胞增殖相关蛋白PAX7表达也较熊果酸组减低。为进一步验证熊果酸靶向自噬调控C2C12成肌细胞增殖,本研究通过流式细胞术检测细胞周期分布,发现熊果酸干预后,G1/G0期细胞减少,而3-MA抑制自噬活性后,G1/G0期周期相对增多,表明熊果酸在一定程度上影响细胞周期和细胞的有丝分裂进程,且这一过程与细胞自噬激活有关。P16INK4a是细胞周期G1/S期检查点的负性调节因子,可以阻遏细胞周期进而抑制增殖,熊果酸可以抑制P16蛋白表达,但其抑制作用在3-MA阻断自噬进程后被逆转。
综上所述,本研究发现熊果酸可以促进小鼠肌源细胞C2C12增殖和自噬,通过3-MA抑制自噬活性后,探讨熊果酸对C2C12细胞增殖调控的机制,发现熊果酸靶向自噬促进C2C12细胞的增殖,为骨骼肌萎缩的治疗及预防提供了新的思路。
