小麦促分裂原蛋白激酶基因TaMAPKK3介导植物抵御低磷功能研究
2020-11-09宋文腾马自飞史美华白菁华路文静
宋文腾,马自飞,史美华,白菁华,路文静,肖 凯
(1.河北农业大学 生命科学学院,河北 保定 071001;2.河北农业大学 农学院,河北 保定 071001 )
促分裂原活化蛋白激酶 (Mitogen-activated protein kinase,MAPK) 级联途径 (MAPKKK-MAPKK-MAPK) 是真核生物中保守的信号传导模块[1],在该级联通路中,通过MAPKKK-MAPKK-MAPK逐级磷酸化的传递方式传递胞内信号[2]。植株体内MAPK级联途径通过将特定信号放大及调控转录因子和细胞骨架的活性等[3],对植株生长、发育和抵御环境逆境的能力产生重要影响[4-5]。模式植物拟南芥中MAPKK,依据编码蛋白的氨基酸序列保守性特征,将该基因家族成员进一步划分为A、B、C、D 4个亚组[5]。其中,A亚组包括AtMKK1、AtMKK2和AtMKK63个基因,B亚组包括AtMKK31个基因,C亚组包括AtMKK4、AtMKK52个基因,D亚组包括AtMKK7、AtMKK8、AtMKK9和AtMKK104个基因。MAPK级联途径中MAPKK蛋白在感受上游信号、激活下游蛋白及传递信号中发挥重要作用[6]。
MAPK特定级联组分基因在介导植株生长发育、免疫响应以及适应各种生物和非生物胁迫过程中发挥着重要调控作用[6-7]。相关研究多以模式植物拟南芥MAPKKK和MAPK家族成员为对象,有关MAPKK家族成员的功能鉴定研究报道尚少[8]。Xu等[8]对植物种属特定MAPKK家族基因介导植株生长和发育的信号传递机制进行了研究。Wersch等[9]对拟南芥MAPKK6介导花青素合成的生物学功能进行了探讨。进一步揭示植物种属MAPKK家族基因特征及信号感受和传递机制,对于深入阐明植物抵御非生物逆境的分子机理具有重要的理论和实践意义。
磷素是作物生长必需的矿质元素,提高作物磷素利用效率对于促进我国农业可持续发展具有重要实践意义[10]。作者在前期开展小麦MAPK级联途径基因的分子特征和表达模式研究中发现,该种属中MAPKK家族成员TaMAPKK3呈低磷逆境下上调表达特征,表明该基因通过转录应答,在介导植株响应低磷逆境中发挥重要作用。基于此,本研究针对迄今有关麦类种属MAPK级联途径分子机制研究有待深入的现状,对该基因的分子特征及介导植株抵御低磷逆境的功能进行了研究,旨在为今后小麦磷高效遗传改良提供理论依据。
1 材料和方法
1.1 试验材料
选用河北农业大学农学院小麦抗逆生理和分子实验室前期鉴定的磷高效普通小麦品种石新828为基因克隆和表达材料,烟草品种Wisconsin 35为转基因受体材料;遗传转化选用大肠杆菌感受态细胞DH5α和农杆菌感受态细胞EHA105;双元表达载体为pCAMBIA3301。以上材料均由小麦抗逆生理和分子课题组所属实验室提供。
试验主要试剂包括RT-PCR试剂、qRT-PCR试剂、限制性内切酶、pUCm-T载体、T4连接酶等,均购自上海生工。
1.2 TaMAPKK3基因克隆及分子特征分析
利用DNAStar工具鉴定供试小麦种属MAPKK家族基因TaMAPKK3(登录号:KT187393)编码蛋白、蛋白分子量和等电点。利用NCBI(https://www.ncbi.nlm.nih.gov/) Blast工具查找供试基因编码蛋白的植物种属同源蛋白,构建系统进化树。
1.3 TaMAPKK3应答低磷逆境的表达模式研究
以前期鉴定的磷高效小麦品种石新828为材料,采用Sun等[11]的方法建立MS溶液培养体系,设置低磷逆境处理小麦幼苗。采用Guo等[12]的方法,鉴定供试基因TaMAPKK3应答低磷逆境下不同时间点的表达模式。扩增目标基因正向引物为:5′-GCTGTTCATTATTACCT-3′;反向引物为:5′-CACCTTTCAACTTCTTT-3′。以小麦组成型表达基因Tatubulin(登录号:U76558)为内参基因(正向引物5′-CATGCTTATCCCTCGTCTCGACCT-3′和反向引物5′-CGCACTTCATGATGGAGTTGTAT-3′)。qRT-PCR反应条件:95 ℃预变性3 min;95 ℃ 30 s,57 ℃ 15 s,循环25次;72 ℃15 s。依据2-ΔΔCT公式计算供试基因各处理时间点的表达丰度。
1.4 低磷处理下TaMAPKK3转化株系植株生长鉴定
采用RT-PCR扩增TaMAPKK3编码阅读框(ORF)。采用的正向引物为5′-AAACCATGGCGGGGCT GGAGGAG-3′(NcoⅠ);反向引物为5′-AAAGGTCACCTCATGCTTGGATAATGTA-3′(BstpⅠ)。将扩增产物融合至表达载体pCAMBIA3301启动子下游,采用Guo等[12]农杆菌介导遗传转化法转化烟草 (cv. Wisconsin 35)。
以获得的T32个典型供试基因转化株系 (Sen-2和Sen-3) 和野生型 (Wild type,WT) 对照为材料,采用前述的小麦低磷处理方法对转化株系和WT进行低磷处理。处理42 d,用相机拍照记录植株表型,采用常规方法测定植株地上组织及根系鲜质量和干质量。
1.5 低磷处理下TaMAPKK3转化株系含磷量和光合参数测定
以低磷处理下上述转化株系和WT为材料,采用Sun等[11]的磷钼黄比色法测定植株地上组织和根系含磷量。通过含磷量与植株干质量乘积获得植株地上组织、根系及植株磷累积量。采用Guo等[12]的方法,利用便携式Li-6400光合系统仪,测定供试材料不同叶位 (第4叶和第6叶) 光合速率 (Pn)、气孔导度 (Gs)、光系统Ⅱ光化学效率 (ΦPSⅡ) 和叶绿素非光化学淬灭系数 (NPQ)。
1.6 数据统计学分析
采用Microsoft Excel 2013和SPSS 10.0软件进行数据录入、处理和统计分析。各测定数据均源于3次测试重复结果。
2 结果与分析
2.1 TaMAPKK3分子特征
TaMAPKK3cDNA全长为1 572 bp,编码523个氨基酸,蛋白分子量58.508 ku,等电点(PI) 5.72。编码蛋白含MAPKKs结构域(78-347 aa)和核转移因子结构域(370-520 aa) 2个结构域, MAPKK结构域内含有激活位点、ATP结合位点和多肽结合位点 (图1-A),属于MAPKKs中B亚组。在氨基酸水平上,TaMAPKK3与乌拉尔图小麦(T.urartu)TuMAPKK6呈序列相似特征 (图1-B),表明它们具有相似的进化途径。

图1 TaMAPKK3蛋白结构域和系统进化特征Fig.1 The domain and phylogenetic properties of the TaMAPKK3 protein
2.2 TaMAPKK3低磷逆境下的表达模式
正常生长条件下,TaMAPKK3的表达水平较低。低磷处理后,该基因表达丰度表现为27 h内随着处理进程不断上升;进行恢复处理后,植株根叶中该基因的表达水平随恢复进程不断降低 (图2)。表明TaMAPKK3呈典型的低磷应答表达模式。
2.3 低磷处理下超表达TaMAPKK3株系的生长特征
采用qRT-PCR对转化株系靶基因转录本进行检测,以显著增多转录本株系Sen-2和Sen-3 (图3-A) 为材料,对其低磷逆境下植株生长特性进行了研究。结果表明,与野生型相比,Sen-2和Sen-3株系植株个体增大、长势更好,根系发育明显改善 (图3-B),植株地上组织和根系鲜质量显著增加,干物质积累量显著提高 (图3-C、D)。上述结果表明,超表达TaMAPKK3基因具有增强低磷逆境下植株生长和改善根系发育的能力。

R.恢复处理。不同小写字母表示差异显著(P<0.05)。图3-4同。 R. Indicates recovery treatment. Different lowercase letters indicate significant difference(P<0.05). The same as Fig.3-4.
2.4 低磷处理下超表达TaMAPKK3株系磷素吸收和光合特性
低磷处理下,与野生型相比,Sen-2和Sen-3植株地上组织和根系磷浓度显著增高 (图4-A),上述器官磷累积量和植株磷总累积量显著增多 (图4-B)。表明TaMAPKK3介导植株抵御低磷逆境能力的增强,与其增强植株根系建成及改善植株磷素吸收能力有关。与野生型相比,转化株系各测试叶位 (第4、6叶) 光合速率 (Pn) (图4-C)、气孔导度 (Gs) (图4-D) 和光系统Ⅱ光化学效率 (ΦPS Ⅱ) (图4-E) 显著提高,叶绿素非光化学猝灭系数 (NPQ)显著降低 (图4-F)。表明TaMAPKK3通过改善根系发育和磷素吸收,促进光合器官碳同化功能,进而增强逆境胁迫下植株干物质生产能力。

图3 低磷处理下TaMAPKK3转化株系生长特征Fig.3 The plant growth characteristic of the TaMAPKK3 overexpressing lines under low-P treatment
3 讨论与结论
MAPK级联途径通过逐级磷酸化进行胞内信号传递的方式,与其通路组分特定蛋白保守位点有关,例如ATP结合位点、激酶活化位点和肽链互作位点等[13-14]。本研究表明,TaMAPKK3编码蛋白含有典型MAPKKs家族蛋白结构域,结构域内存在激酶活化位点、ATP结合位点等功能位点;在370-520 aa组成NTF2结构域,表明TaMAPKK3属于B亚组MAPKKs,有关上述结构域的功能有待进一步研究。本研究发现,TaMAPKK3在进化关系上与乌拉尔图小麦TuMAPKK6蛋白高度同源,说明它们可能源自同一祖先,在介导信号传递中发挥相似的生物学功能。
特定MAPK级联通路组分家族基因,会通过提高转录丰度参与对特定逆境的响应[15-16]。例如,当拟南芥遭遇盐胁迫时其MPK4和MPK6表达量明显上升[17-18]。在本研究中,在低磷处理下,TaMAPKK3转录丰度随处理时间的加长提升,复磷处理后,供试基因转录本恢复到初始水平。表明该基因通过在转录水平上对低磷逆境信号产生应答,进而参与对低磷逆境信号的转导过程。有关该小麦MAPKK家族基因应答低磷逆境的启动子顺式作用元件及转录机制有待进一步探讨。
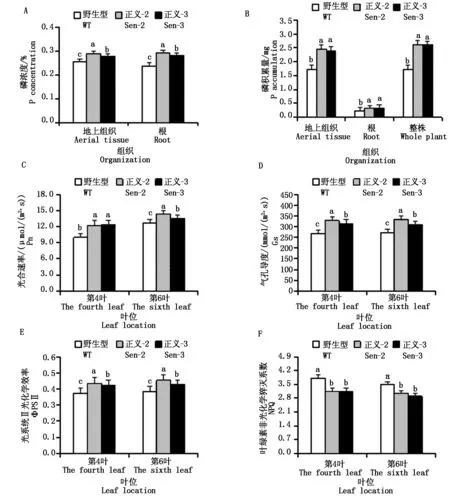
图4 低磷处理下转化株系的磷素吸收和光合特性Fig.4 The Pi uptake and the photosynthetic characteristic of the transgenic lines under low-P treatment
异源表达拟南芥AtMPKK2烟草株系,低温和盐分耐受性显著增强[19-20]。本研究通过农杆菌介导法获得超表达TaMAPKK3的转基因烟草植株;低磷处理下,与野生型相比,超表达TaMAPKK3烟草植株生长状况显著改善,植株磷含量增加,光合碳同化能力增强。说明TaMAPKK3能够提高烟草植株抵御低磷胁迫的能力。有关TaMAPKK3增强植株耐低磷胁迫的分子机制尚有待进一步研究。
TaMAPKK3编码蛋白含有MAPKKs结构域和NTF2蛋白结构域,归属于MAPKKs B亚组,在氨基酸水平上与乌拉尔图小麦TuMAPKK6呈高度序列一致性特征。该基因呈典型低磷诱导表达模式。低磷处理下,超表达TaMAPKK3烟草植株较野生型长势增强,干质量增多,磷素积累量增加,光合能力提高。综合说明TaMAPKK3在介导植株抵御低磷胁迫中发挥重要作用。
