丛枝菌根真菌与有机肥配施对甜玉米根际土壤关键碳循环功能基因的影响
2020-10-31袁银龙徐如玉左明雪顾文杰卢钰升解开治徐培智
袁银龙,孙 杰,徐如玉,左明雪,顾文杰,卢钰升,解开治,徐培智
(1.甘肃农业大学资源与环境学院,甘肃 兰州 730070;2.广东省农业科学院农业资源与环境研究所,广东 广州 510640)
0 引言
【研究意义】碳是有机体组成的必需元素,同时碳循环也是地球生物化学循环中重要的循环过程之一。微生物在碳循环过程中起着不可替代的作用,介导了包括碳分解、碳固定、甲烷代谢等3 个碳循环的基本过程[1-2]。而农田土壤碳循环对农田土壤肥力的形成与维持、农田生产力的提高有着重要意义。土壤微生物是土壤碳循环的主要驱动力,在土壤有机碳转化和养分循环方面起着关键性作用[3]。因此,农田土壤碳转化的研究显得日益重要,已逐渐成为国内外研究的热点和重点[4]。丛枝菌根真菌(Arbuscular mycorrhizal fungi, AMF)能促进植物对多种营养元素的吸收和利用,并调节农田生态系统养分循环[5]。针对我国南方甜玉米栽培化肥施用量大、利用效率低的突出问题[6-7],在大田条件下研究AMF 与有机肥配施对甜玉米根际土壤微生物碳循环功能基因的影响,明晰其对甜玉米-土壤碳循环的微生物学过程尤为重要。【前人研究进展】田善义等[8]认为有机无机肥配施能不同程度促进参与碳循环的总体酶活性的增加,在满足微生物和植物养分的同时可以提高土壤生物活性和生态代谢效率。前人研究表明[9],菌根真菌不但具有重要的土壤碳汇功能,同时菌根真菌也可参与土壤复杂有机质的降解过程。金海如等[10]研究发现不同有机物物料与根内球囊霉配施,有利于增加玉米籽粒中磷素和氮素含量,也提高了可溶性糖、淀粉、蛋白质和氨基酸等含量。徐如玉等[11]发现对不同施氮比例下增施摩西管柄囊霉(Funneliformis mosseae)可显著提高甜玉米的氮肥利用效率,改变AM 真菌群落结构,提高甜玉米产量,是甜玉米氮肥减量的有效途径。徐丽娇等[12]研究表明,低磷胁迫下AM真菌可能通过分泌物调控植物碳磷代谢相关基因的表达,进而调节植物对低磷胁迫的生理响应。目前,对于农田土壤碳循环的研究多使用Biolog平板培养法,如通过对烤烟[13]、转基因玉米[14]、燕麦[15]等根际土壤微生物利用不同碳源的能力来分析其碳代谢的能力。【本研究切入点】目前,关于利用功能基因芯片(GeoChip5.0)技术对碳循环的研究多集中在冻土、高寒草甸、深海等生态系统,对于农田土壤碳循环的研究多使用Biolog 平板培养法。在农田土壤碳循环中,尤其是利用GeoChip 对AMF配施有机肥下甜玉米根际土壤碳循环的研究甚少。【拟解决的关键问题】本研究采用大田区组设计,利用功能基因芯片(GeoChip5.0)技术,研究AMF配施有机肥对甜玉米根际土壤碳循环微生物功能基因的影响,明晰甜玉米-土壤碳循环的微生物学过程,以期为甜玉米化肥减施增效提供技术支撑。
1 材料与方法
1.1 试验地概况及试验材料
试验地位于广东省惠州市惠阳区农业技术推广中心试验基地(23°8′46.19″N,113°20′47.84″E),于2018 年春季开始,1 年春秋2 季栽培。试验区气候类型为亚热带季风气候,年平均气温为13~22 ℃,年平均降水量800~1 500 mm,土壤类型为砖红壤。试验开始前基本地力养分参数为有机质含量22.8 g·kg-1,碱解氮191.54 mg·kg-1,有效磷49.56 mg·kg-1,速效钾149.41 mg·kg-1,pH 值5.18。供试甜玉米(Zea Mays L.)品种为粤甜28 号,由广东省农业科学院作物研究所提供。供试菌种为变形球囊霉菌(Glomuse versiforme, BGC GD01C),购自北京市农林科学院植物营养与资源研究所。变形球囊霉(Glomuse versiforme)菌剂含被侵染的甜玉米植物根段、菌根真菌孢子及根外菌丝的根际沙土混合物,其孢子密度为38 个·g-1。孢子的扩繁参考孙丹萍等[16]和毕银丽等[17]的方法进行室内盆栽砂土扩繁,具体操作同参考文献[11]。
1.2 试验设计
试验共设置7 个处理,每个处理设置3 个重复,共计21 个小区,各小区面积54 m2(4.5 m×12 m),随机区组排列,甜玉米种植密度为45 000 株·hm-2,起垄种植。根据当地高产栽培方式优化最佳施肥量为:氮肥(N)330 kg·hm-2、磷肥(以P2O5计)150 kg·hm-2、钾肥(以K2O 计)225 kg·hm-2,具体施肥方案如表1。
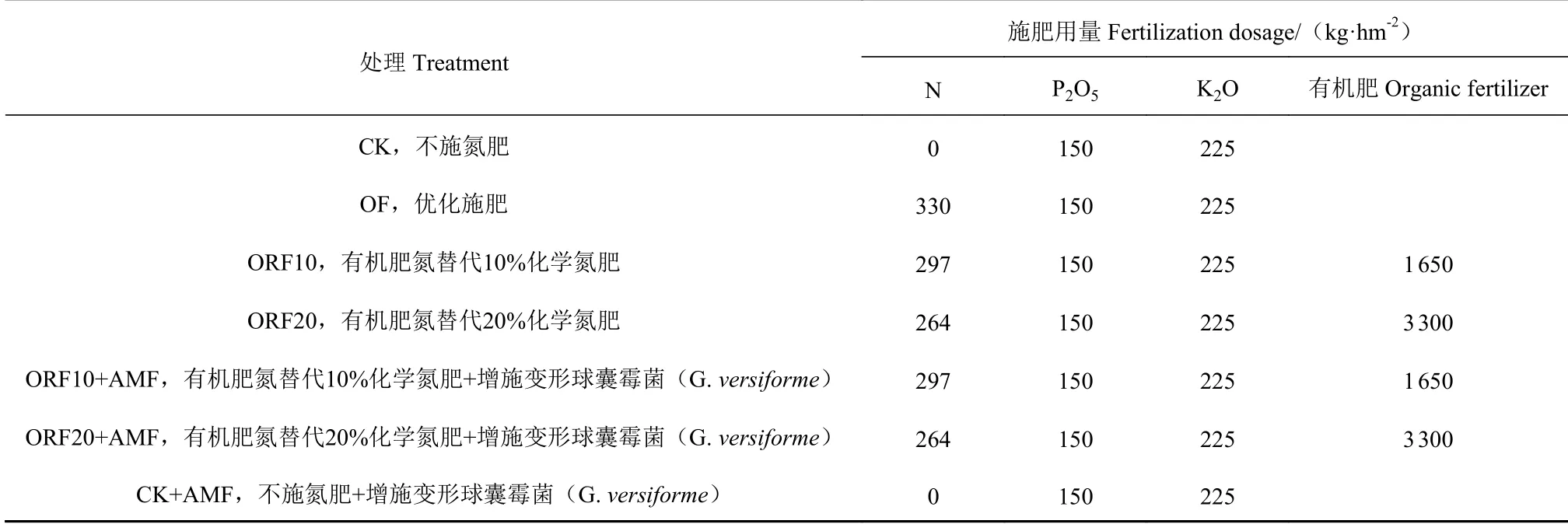
表1 不同处理施肥方案Table 1 Fertilization treatments
试验于2019 年9 月18 号进行幼苗移栽,甜玉米幼苗移栽时增施变形球囊霉菌菌剂,每株定植穴接种7 g;未接种菌剂的处理,每株接种无菌基质7 g。
氮肥使用尿素,在基肥、苗期、拔节期分别以2∶3∶5 的比例施入;磷肥使用过磷酸钙(P2O512%)一次性基施,钾肥使用硫酸钾(K2O 50%),在基肥和拔节期分别以5∶5 的比例施入。商品有机肥为广州新农科肥业科技有限公司生产,主要以优质堆肥原料牛粪、蘑菇渣、米糠等经过高温堆肥制备而成。经测定,其养分含量分别为N 2%、P(P2O5)1.5%、K(K2O)1.5%。有机肥作为基肥一次性施入,施肥量为商品有机肥产品质量,其他田间管理按照当地田间管理模式进行。
1.3 根际土壤采集方法
土壤样品于成熟收获期采集,每个小区选取5 株具有代表性的甜玉米植株将其整棵挖出,参照Zhao 等[18]的抖根法获取甜玉米根际土壤约1 500 g,然后将土壤混合均匀分为2 份,一份室温保存带回风干用于理化性质的测定,一份放入无菌密封袋中用冰盒带回用于甜玉米根际土壤微生物群落多样性及碳循环功能基因的分析。
1.4 土壤理化性质测定方法
土壤pH 值采用酸度计;全氮采用凯氏定氮法;全磷和全钾经消煮后,采用钼锑抗比色法和火焰光度法;有机质采用重铬酸钾容量法;土壤速效磷采用氟化铵浸提(酸性土)钼锑抗比色法;速效钾采用乙酸铵浸提火焰光度法测定。以上土壤常规理化性质均参照鲍士旦土壤农化分析方法测定[19];土壤呼吸采用室内密闭培养法等进行测定[20]。
1.5 甜玉米鲜苞测产
供试甜玉米鲜苞于2019 年秋季甜玉米生长的成熟期(12 月10 日)采集,各小区全部实收测产并折算为每公顷产量。
1.6 功能基因芯片分析
1.6.1 土壤DNA 的提取 DNA 提取和定量基因组DNA 由广东美格基因生物技术有限公司(中国广州)使用商业试剂盒(MinkaGene Soil DNA Kit),根据制造商的操作流程提取。DNA 质量由NanoDrop One(Thermo Fisher Scientific, Waltham, USA)评估,最后DNA 浓度通过Pico Green 使用FLUOstar Optima 微板读取器(BMG Labtech,Jena,Germany)进行量化。
1.6.2 DNA 标记杂交 用随机引物和DNA 聚合酶I 的Klenow 片段对纯化后的DNA 进行Cy3 染色[21]。用QIA 快速纯化试剂盒(Qiagen,Valencia,CA,USA)纯化标记的DNA,然后在Labconco 离心浓缩器(Labconco Corp,Kansas City,MO)中50 ℃干燥45 min。干燥后的DNA 稀释至相同浓度,然后在95 ℃孵育5 min,并保持在42 ℃进行杂交16 h。随后,标记的DNA 被放置到数组,杂交的数组预热到42 ℃(MAUI,BioMicro Systems,Salt Lake City,UT,USA)至少5 min。最后,微阵列是由罗氏MS200 扫描仪扫描(Roche,Madison,WI,USA)获取光信号。
1.6.3 数据的处理与标准化 利用ImaGene 6.0(Biodiscovery Inc.El Segundo,CA,USA)软件将光信号转换为数字信号,得到探针信号强度。去除信噪比(SNR)<2 的斑点。如果在至少2·3-1的复制集中检测到探针的信号,则认为探针为阳性。数据归一化基于对数变换,确定每个样本的平均信号强度,然后将每个探头的强度按对应样本的平均强度进行放大与归一化处理,得到标准化后的标准数据。
1.7 数据分析与处理
图表统计分析采用Excel 2013 和SPSS 24.0,GraphPad Prism 8 进行作图,施菌处理和有机无机配施水平及其交互作用的差异显著性采用双因素方差分析(Two-way ANOVA)进行检验(P<0.05);检验不同处理之间的差异显著性采用Duncan’s 多重检验法进行检验(P<0.05);DCA 与RDA 分析和绘图采用QIIME2 和ggplot2 包。
2 结果与分析
2.1 AMF 配施有机肥对甜玉米产量、土壤有机质和土壤呼吸的影响
由表2 可见,在不施氮的情况下,单施菌肥的玉米鲜苞产量显著大于对照处理(P<0.05);不同有机肥替代水平与优化施肥的鲜苞产量无显著差异,但与对照的差异比较明显(P<0.05),在有机肥替代水平上增施变形球囊霉菌后,发现同替代水平下均为施菌产量大于不施菌产量(P<0.05),但同样施菌水平下,不同替代水平的甜玉米鲜苞产量没有显著差异。在不施氮肥、有机肥替代10%化学氮肥、有机肥替代20%化学氮肥水平下,施用菌肥较不施用菌肥的鲜苞产量分别提升32.6%,8.6%、8.9%。有机质与土壤呼吸均表现为有机肥替代20%化学氮肥处理高于其他处理,其中土壤呼吸指标显著高于其他处理(P<0.05)。总体来看,同一有机肥替代水平下,施用菌肥显著提高甜玉米产量。增施变形球囊霉菌和有机无机配施均极显著地影响产量、土壤呼吸,但二者除土壤呼吸外均无显著的交互效应。
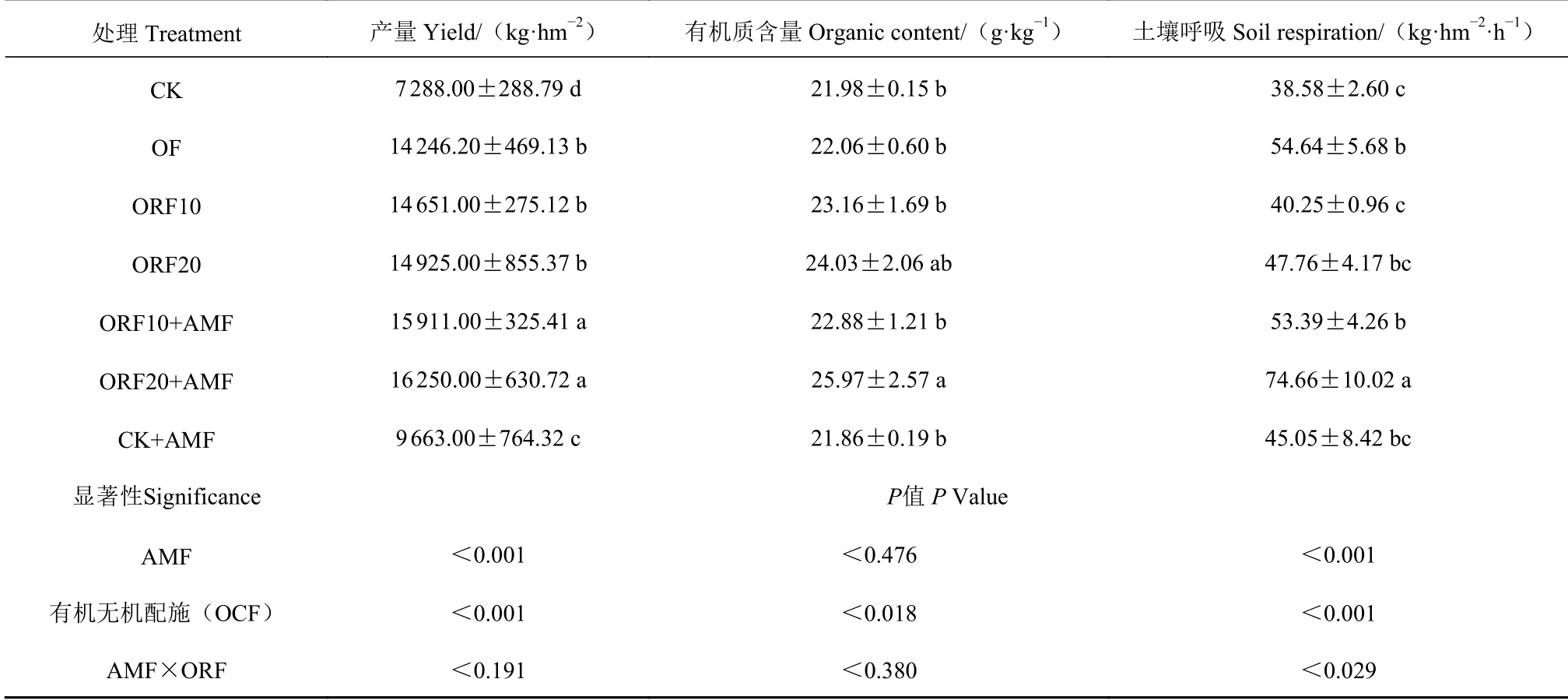
表2 不同施肥下甜玉米产量与理化性质分析Table 2 Yield and physicochemical properties of sweet corn under different treatments
2.2 碳循环功能基因结构特征
对7 个不同施肥处理碳循环基因的结构特征进行DCA 排序分析(图1),可以看出7 个不同施肥处理在排序图中没有重叠,能够清楚地分开,说明不同施肥处理下与碳循环有关的基因的结构特征发生了明显的变化。施用变形球囊霉菌的3 个处理均落在DC2 负半轴,不施菌肥的4 个处理,大多数在DC1 负半轴。相对来说,不施形球囊霉菌的4 个处理在距离上较近,它们的碳循环功能也更为接近。施用AMF 的3 个处理中,ORF20+AMF 与CK+AMF两个处理在DC2 轴上的距离较近,因此ORF20+AMF与CK+AMF 两个处理的关键碳循环功能基因的结构组成接近。其中OF 与ORF20+AMF 处理在DC1 和DC2 轴上均有一定的距离,说明OF 与ORF20+AMF处理关键碳循环功能基因的结构组成差异明显。

图1 碳循环功能基因结构特征Fig.1 Structural characteristics of C-cycle genes
2.3 关键碳循环功能基因的标准化信号强度
21 个样品中共检测到169 911 个与碳循环有关的基因,其中碳分解115 575 个、碳固定49 677 个、甲烷代谢4 659 个。由图2 可以看出,CK+AMF 与ORF20+AMF 处理的碳降解过程中淀粉(Starch)降解基因(amyA, amyx, pula)、半纤维素(Hemicellulose)降 解 基 因(mannanase, xyla, xylanase)、 纤 维 素(Cellulose)降 解 基 因(cellobiase, endoglucanase,exoglucanase)、几丁质(Chitin)降解基因(acetylglucosaminidase, chitinase)的信号强度显著高于其他处理(P<0.05)。淀粉作为最容易被微生物利用的碳源,在被检测到的基因信号强度也是所有碳源中最高的,说明分解利用淀粉的微生物是最活跃也是数量占有优势的。在检测到的19 个与碳分解有关的基因在处理间的差异均达到了显著水平。总体而言,CK+AMF 是所有处理中碳分解基因信号强度最高的处理,ORF20+AMF 次之。
如图3,参与碳固定的关键基因如rusbico、aclb、CODH 和pcc 在21 个样品中均被检测到。在检测到的4 个碳固定的基因信号强度各处理间差异均达到显著水平,全部表现为CK+AMF 最高,ORF20+AMF次之,ORF10 最低。参与甲烷生成的基因mcra、甲烷氧化的基因mmoX、pmoa 等3 个与甲烷代谢有关的基因在21 个样品中均被检测到。在参与甲烷代谢的3 个基因在不同处理间的变化趋势与碳分解和碳固定保持一致,仍然表现为CK+AMF 最高,ORF20+AMF 次之,ORF10 最低,并且参与甲烷代谢的3 个基因在7 个处理中均差异显著(P<0.05)。
2.4 环境因子对微生物群落碳循环功能基因的影响
利用碳循环功能基因与土壤环境因子数据矩阵进行冗余分析,可以反映土壤环境因子与土壤碳循环功能基因之间的关系及相关程度。由图4 所示,图4-a 第一排序轴和第二排序轴分别解释了67.48%和29.84%,图4-b 第一排序轴和第二排序轴分别解释了67.48%和29.84%,都较好地解释了碳循环功能基因强度与土壤环境因子之间的关系。
增施变形球囊霉菌的处理(图4-a)在碳降解过程中,淀粉降解基因(amyA, amyx, apu)与土壤呼吸(SR)、全氮(TN)、有机质(OM)、速效磷(AP)和全磷(TP)呈正相关;半纤维素降解基因(ara,mannanase, xylose-isomerase-Oomycetes)与全氮、有机质呈正相关;纤维素降解基因(endoglucanase,exoglucanase)与土壤呼吸、全氮、有机质、速效磷和全磷呈正相关;几丁质降解基因(acetylglucosaminidase, chitinase)与土壤呼吸、全氮、有机质、全磷和速效磷呈正相关;果胶降解基因(RgaE)与速效钾(AK)、全钾(TK)和pH 呈正相关;木质素降解基因(phenol-oxidase)与速效钾、全钾和pH 呈正相关;在碳固定过程中:rubisco、pcc 与全氮、速效磷、全磷和有机质呈正相关;产甲烷(mcra)与土壤呼吸、速效钾、pH 和全氮呈正相关;在甲烷代谢中甲烷氧化的mmox 与全钾、土壤呼吸呈正相关;pmoa 与速效磷、全磷和有机质呈正相关。
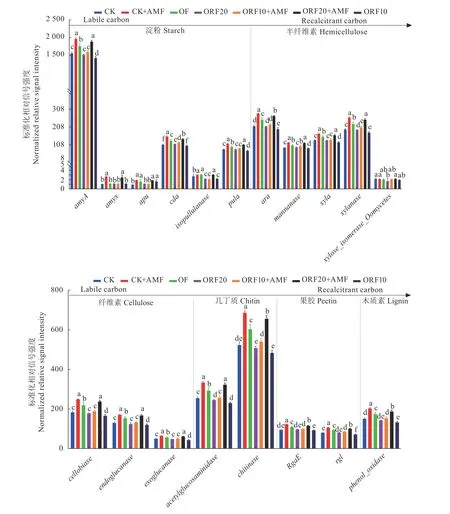
图2 不同处理下碳降解的关键功能基因标准化相对信号强度Fig.2 Normalized relative signal strength of key functional genes relating to C-degradation under different treatments
在不施变形球囊霉菌的处理(图4-b)在碳降解过程中,淀粉降解基因(amyA, cda)与全氮、速效磷、全磷和有机质呈正相关关系,与其他环境因子呈负相关关系;半纤维素降解基因(ara, mannanase,xyla, xylanase)与速效钾、pH、全氮、速效磷、全磷和有机质含量呈正相关,与其他环境因子呈负相关,纤维素降解基因(cellobiase, endoglucanase,exoglucanase)与全钾、速效钾和土壤呼吸呈正相关,几丁质降解基因(acetylglucosaminidase)与全钾、土壤呼吸呈正相关;果胶降解基因(RgaE, rgl)与全氮、速效磷、全磷、有机质呈正相关,与其他环境因子呈负相关;木质素降解基因(glx, ligninase,mnp)与速效钾、pH、全氮、速效磷、全磷和有机质呈正相关,与其他理化因子呈负相关。在碳固定过程中:rubisco、pcc 与全氮、速效磷、全磷和有机质呈正相关,与其他环境因子呈负相关。在甲烷代谢过程中产甲烷(mcra)与土壤呼吸、速效钾、pH 和全氮呈正相关,与其他环境因子呈负相关,甲烷氧化的mmox 基因与全钾、土壤呼吸呈正相关,与其他环境因子呈负相关,pmoa 基因与速磷、全磷和有机质呈正相关,与其他环境因子呈负相关。

图3 不同处理下碳固定与甲烷代谢的关键功能基因标准化相对信号强度Fig.3 Normalized relative signal strength of key functional genes relating to C-fixation and

图4 碳循环基因与理化性质的冗余分析Fig.4 Redundant analysis on carbon cycle genes and physicochemical properties
Monte Carlo 检验显示,在不增施变形球囊霉菌的处理中,土壤呼吸(P=0.024,解释量为74.86%)、全钾(P=0.024,解释量为76.49%)对碳循环功能基因的影响最为显著;在增施变形球囊霉的处理中,速效磷(P=0.002,解释量为88.07%)、pH(P=0.016,解释量为84.69%)、有机质(P=0.004,解释量为83.67%)、土壤呼吸(P=0.008,解释量为80.21%)、全氮(P=0.006,解释量为79.57%)、全磷(P=0.011,解释量为76.33%)对碳循环功能基因的影响最为显著。
3 讨论
近年来,由于片面追求产量而过量施氮的施肥模式不仅造成了资源浪费,还带来了环境污染等一系列问题,也给农业生产带来了重大的经济损失[22]。研究表明,AMF 与有机肥配施能在一定程度上促进作物对养分的吸收利用[5],并且提高作物品质[23-24]。通过AMF 与有机肥配施来调节作物利用养分方式与农田生态系统养分循环,从而达到维持农田生态系统可持续发展与利用率的提高。
前人研究表明,AMF 与有机肥配施对作物有极其显著的增益效果。金海如等[10]在AMF 与有机肥配施的研究中发现,AMF 与有机肥配施能显著提高甜玉米的产量与品质;同样,周世品[24]也发现有机无机肥配施和丛枝菌根化育苗对西瓜产量品质有明显的提高。本研究中,在CK、有机替代10%、有机替代20%施肥水平下,AMF 与有机肥配施较不施用菌肥的鲜苞产量分别提升32.6%、8.6%、8.9%。说明AMF 与有机肥配施能显著提高甜玉米的产量。接种变形球囊霉菌后提高了作物对各种养分的利用能力和效率,促进了甜玉米的生长,从而达到提高甜玉米产量的效果[5]。土壤呼吸是陆地生态系统碳循环中最活跃的部分,能在一定程度上反映土壤的物质代谢强度、生物学特性以及土壤碳库的稳定性[25]。贺美等[26]通过对黑土长期定位的研究发现,有机无机配施能显著提高土壤呼吸的强度,但本研究结果在CK、ORF10、ORF20 等3 个施氮水平上增施变形球囊霉菌后对土壤呼吸强度的提升最为明显。最近的研究结果表明丛枝菌根真菌具有机质降解的功能[27-28],这可能也是ORF20+AMF 处理的土壤呼吸强度高于其他处理的原因。
土壤碳循环是地球生物化学循环过程中最重要,也是最复杂的循环过程之一[29]。本研究检测到的参与碳分解过程的19 个基因的信号强度在各处理间均具有显著差异,碳固定与甲烷代谢的7 个基因的信号强度在处理间具有显著差异。研究结果表明,碳固定过程中,Calvin cycle(卡尔文循环)是二氧化碳固定的最主要途径之一,在陆地生态系统中,卡尔文循环存在于从细菌、真菌、藻类到绿色植物的各种生物体中[30]。在卡尔文循环中,RubisCO催化卡尔文循环中的第一步CO2固定反应,使大气中的CO2转化为有机的细胞成分固定下来[31]。AclB和pcc 分别是编码还原型三羧酸循环与3-羟基丙酸双循环的功能基因[32]。本研究中4 种碳固定基因信号强度均表现为CK+AMF、ORF20+AMF 两个处理大于其他处理,说明CK+AMF、ORF20+AMF 处理较其他处理有更强的固碳能力。Gifford 等[33]认为陆地生态系统可以通过固碳来减缓CO2的增加,从而减缓气候变化。但Van Groenigen 等[34]认为陆地生态系统固碳的能力被高估了。同样,在易降解碳到难降解碳的6 种碳源分解中,19 个与之相关的基因也表现出相同的趋势,CK+AMF、ORF20+AMF 的基因信号强度总体上高于其他处理。Xue 等[35]研究表明,土壤中的有机碳含量随着碳降解基因的增加而出现降低的趋势,这说明碳降解基因丰度的增加能提高微生物对碳源的分解能力。土壤微生物介导的农田土壤碳循环体现在温室气体的产生和温室气体的消耗两个相反的方面,温室气体的生成包括CO2和CH4的生成,温室气体的消耗则包括CO2的固定和CH4的氧化[4]。部分学者[36-37]认为增加的CO2也会刺激土壤N2O 和CH4的排放,而这些气体的全球变暖潜力远远高于二氧化碳:N2O 比CO2高298 倍,CH4比CO2高25 倍。He 等[38]利 用GeoChip 技术对雪松湾自然保护区的土壤微生物在CO2升高下对固碳基因pcc 的影响进行研究,结果表明随着CO2浓度的增加,与固碳有关的pcc 和RubisCO 基因的信号强度也随之增加。本研究中,AMF 与有机肥配施后CK+AMF、ORF20+AMF 处理无论是碳固定还是碳降解有关的基因相对信号强度都显著增加,说明在碳分解与固定能力上有了较大的提高。这可能会短期分解碳源增加,促进植物生长。此外,本研究中碳固定基因丰度的增加有可能是碳分解产生更多的CO2刺激下的结果。
在以往利用GeoChip 的研究表明,检测到的功能基因信号强度与环境养分含量有显著的相关性,可以在一定程度上将微生物群落与生态系统过程和功能联系起来[39-40]。前人研究认为植物的多样性与土壤养分是影响微生物功能结构的重要环境因子[41-42]。本研究冗余分析显示增施变形球囊霉菌和不增施裂区均解释了97.32%的碳循环功能基因与环境因子之间的关系,说明所选的环境因子较好地解释了碳循环功能基因与环境因子之间的相关程度。其中土壤呼吸、全钾、速效磷等多种环境因子是影响碳循环功能基因的主要环境因素。Zhang 等[43]通过对高寒草甸微生物碳代谢功能基因的研究表明,全氮、全磷、pH 等土壤养分是影响微生物结构和功能的主要因素。在施用变形球囊霉菌的处理中,pH以84.69%的解释量与碳循环显著相关。Rousk 等[44]也认为土壤pH 是影响微生物功能的关键因子。土壤呼吸作为土壤碳循环过程的间接评估指标,与碳循环有密不可分的关系[45-46]。在本研究中土壤呼吸无论是在施用变形球囊霉菌还是不施用变形球囊霉菌的处理中,都与碳循环基因具有显著相关性。
4 结论
接种变形球囊霉菌配施有机肥与未接种变形球囊霉菌配施有机肥的处理相比显著提高了土壤呼吸强度和甜玉米鲜苞产量;ORF20+AMF 处理在减施20%化学氮肥下甜玉米鲜苞产量显著高于OF 处理,可作为推荐施肥方案。
接种变形球囊霉菌配施有机肥显著增强碳降解过程中淀粉降解基因(amyA,amyx,pula)、半纤维素降解基因(mannanase,xyla,xylanase)、纤维素降解基因(cellobiase,endoglucanase,exoglucanase)、几丁质降解基因(acetylglucosaminidase)的强度。
接种变形球囊霉菌配施有机肥可显著提高碳固定基因(rubisco,pcc,aclb)、甲烷代谢基因(mcra,mmox,pmoa)的相对信号强度。
