广西陆川不同树种人工林的薇甘菊入侵程度变化
2020-10-31黄小荣曹艳云何燚陆国导曹纪文刘庐
黄小荣,曹艳云,*,何燚,陆国导,曹纪文,刘庐
1. 广西壮族自治区林业科学研究院,南宁 530002
2. 广西陆川县林场,广西陆川 537716
0 前言
薇甘菊(Mikania micrantha)为菊科假泽兰属多年生藤本植物,原产中南美洲,于20世纪80年代初期传入香港和华南地区[1]。在万方浩等[2]的统计中,中国大陆的薇甘菊仅分布于广东省;近年来,薇甘菊迅速传播至云南、贵州、福建、广西[3]。薇甘菊2003年进入广西梧州[4],2008年出现在陆川县铁路沿线[5],2010年在陆川县乌石镇巨尾桉林边被发现[6],至 2018年 1月广西区内薇甘菊危害面积已达 5227 hm2;广西受薇甘菊危害最为严重的地区就是公路旁桉树林[7];薇甘菊通过攀援、覆盖、捆绑桉树,严重阻碍桉树生长。广西是桉树大省,桉树是广西近十年来土山造林的最主要树种,2018年全区桉树面积达213万公顷。陆川县目前 80%的林业用地种植桉树;近几年来,部分群众试图采用更换造林树种和密植等方法来减轻薇甘菊对人工林生长的危害,如改种金合欢属植物等。对于薇甘菊的生物学和生态学前人进行了相当密集的研究,如黄忠良等[8]发现薇甘菊喜光;弱光可能抑制薇甘菊光系统功能的形成[9];林下光照是幼苗生长和生存的主要影响因子之一[10];薇甘菊种子生产[11]、种子萌发[12]都需要较充足的光照。这些研究多是在广东和云南省开展的,且大多数是在植物园、自然保护区、农用地和实验室进行;缺乏山地人工林的调查研究,欠缺将薇甘菊与桉树特性相联系的延展思考。
探索薇甘菊入侵与环境因子关联性对薇甘菊防治和疫区合理选择造林树种具有重要意义。本研究在广西陆川薇甘菊严重危害区对相思、桉树人工林进行了实地调查,拟探讨以下问题: 哪些环境因子显著影响人工林中薇甘菊盖度和开花数量?薇甘菊疫区还适合种植桉树吗?
1 研究方法
1.1 研究地点
研究区位于广西陆川县乌石镇王沙村,距桂粤省界 5 km 左右;地理位置北纬 22°6.07′,东经110°17.58′,最高的样地海拔 158 m,最低海拔 98 m,低山丘陵地貌,山坳冲沟有小溪流汇入清湖水库。王沙村水泥乡道较多,多沿山脚蜿蜒,无隧道开凿或劈山开路等重度人为干扰。研究区近热带海洋,属南亚热带季风气候;土层深厚肥沃,雨量充沛,年平均降雨量为 1900 mm;年平均温度为 21.7℃,热量丰富,阳光充足。
1.2 样地概况
在王沙村东南部 3个地点进行样方调查,编号为生境1、生境2、生境3,生境间相距336-500 m,3个生境包括杂交相思(Acacia mangium×Acacia auriculiformis)、火力楠(Michelia macclurei)、巨尾桉(Eucalyptus grandis×E.urophylla)和马占相思(Acacia mangium)4个树种人工林的6个林分;分别由陆川县林场、村民、广西林科院和高峰林场营建,株行距均为 2.5 m × 2 m,详情见表 1。

表1 不同生境6个林分的基本情况Table 1 Profiles of 6 stands at investigated sites
1.3 调查方法
样方设置参考Elzinga等[13]的系统抽样法,有改动。以山脚林缘为基线,与基线垂直向山上拉截线,截线上间隔10 m取1个1 m × 1m小样方记录林下植物名,以小样方为中心的直径5 m样圆记录样方环境因子,包括上层林冠盖度、林下植被盖度、薇甘菊盖度、物种丰富度、坡度、流径长(Flow path length)、距林缘距离、朝向等。野外调查中分层记录植被结构[14],上层林冠盖度是3 m以上层次植物枝叶的垂直投影在样圆中所占百分比,林下植被盖度是3 m以下层次植被盖度;林冠郁闭度通常指维管植物的垂直投影[15],本文分上下两层来分析植被盖度对薇甘菊盖度的影响。流径长根据运动路径算法(Kinematic Routing Algorithm)粗略目测[16],流径长表征地形湿润程度;朝向参考王孝安[17]介绍的方法进行了转换:式中,aspect为朝向的方位角度,转换后的朝向值TRASP由小到大表示生境越来越干热;每个生境的调查涵盖面积均大于2 hm2。调查时间在2018年11月中旬,正值薇甘菊盛花期。生境 1的样方记录了薇甘菊有花、无花情况;有花状况又细分为“微开、开、盛开”三级,分别对应薇甘菊复伞房花序 1—2个、3—6个、7个及以上。
1.4 统计分析方法
用 SPSS 24.0进行多重比较,方差齐性时用LSD,方差非齐性用T2法。薇甘菊盖度对环境因子变化的响应采用线性混合效应模型,使用 R 3.5.2[18]的nlme包,模型中均以薇甘菊盖度为响应变量、林分为随机截距,解释变量有多种组合,以下简称“混合效应模型”,作图和分析参考 Zuur[19],互作效应分析采用effects包[20]。薇甘菊是否开花的二元目标变量协方差分析使用 R 软件的广义线性模型,指定误差分布为二项式[21],协方差模型包括环境因子的主效应和两两互作;参考黄小荣等[22]的方法,建模后逐步减掉不显著的互作项、主效应以得到最终模型。
2 结果与分析
2.1 上层林冠盖度、林下植被盖度的多重比较
6个林分的上层林冠盖度均值比较差异极显著(P< 0.000),降序为 B、F、A、C、D、E;林分 C 即3.5 yr桉树人工林的上层林冠盖度特别低;林分 D和 E的上层林冠盖度接近零是因为林龄短;除桉树林外,其他 5个林分均基本符合林龄越大上层林冠盖度越高的规律(表2)。林下植被盖度在6个林分之间差异极显著(P< 0.000);3.5 yr桉树林的林下植被盖度很高(95.8%),火力楠的林下植被盖度极低(8.3%),林分A杂交相思和林分E桉相混交的林下植被盖度中等(表2)。
2.2 薇甘菊盖度的均值比较
不同生境的薇甘菊盖度均值差异极显著(P <0.000),生境1、2、3薇甘菊盖度均值分别为30.9%、10.4%和13.6%,在生境1的林缘,薇甘菊密如蛛网,繁殖体压力很大。6个林分之间比较薇甘菊盖度差异极显著(P= 0.000),林分A杂交相思的薇甘菊盖度最高(34.2%),林分 B火力楠的薇甘菊盖度为零(表2)。上层林冠盖度变量分箱成≤20%、21%—60%和>60%三个等级,三个等级的薇甘菊盖度差异极显著(P= 0.001),均值分别为 16.4%、36.1%和 12.0%,LSD多重比较结果为B、A、B,上层林冠盖度中等的样方薇甘菊盖度极显著较高(P< 0.01)。
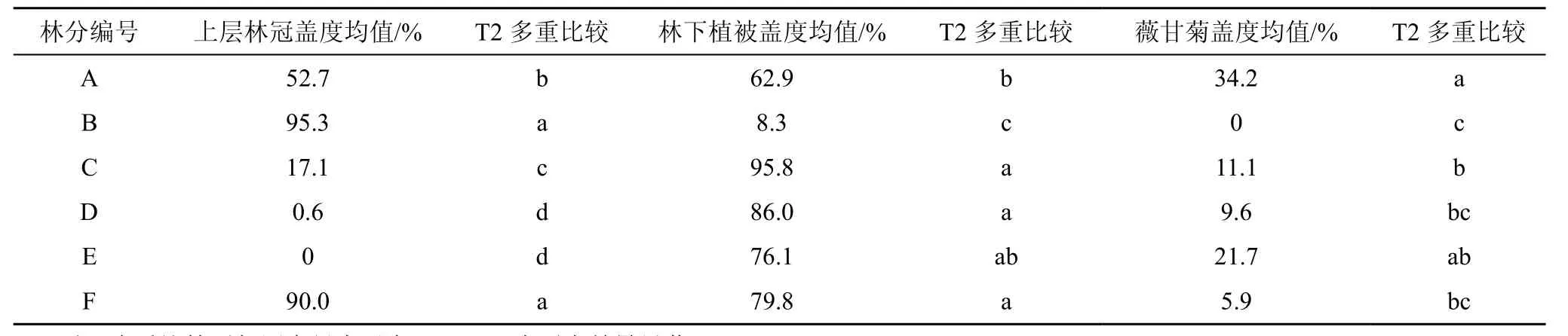
表2 不同林分的植被盖度和薇甘菊盖度多重比较Table 2 Multiple comparisons of vegetation and Mikania micrantha cover against stands
2.3 薇甘菊盖度对环境因子变化的响应
以薇甘菊盖度为响应变量、林分因子为随机截距、其他所有环境因子为解释变量建立线性混合效应模型,经过简化后只剩林下植被盖度一个因子显著(P< 0.000),薇甘菊盖度随林下植被盖度增加而增加。分别以单个因子为解释变量的混合效应模型分析发现,上层林冠盖度和朝向值TRASP对薇甘菊的单因素效应显著,P值分别为0.027和0.028,两个因子均与薇甘菊盖度负相关;因此共有 3个单因素主效应显著影响薇甘菊盖度(表3)。
单因子效应显著的 3个因子的两两互作中,上层林冠盖度和 TRASP对薇甘菊盖度的互作效应显著(P= 0.019),其他两对互作效应不显著;在较湿冷立地,薇甘菊盖度随上层林冠盖度增加微弱减少;在较干热立地,薇甘菊盖度随上层林冠盖度增加急剧减少。
混合效应模型允许每个林分有不同的截距;比如上层林冠盖度的混合效应模型中,林分A、B、C、D、E和 F的截距分别为 45.3、23.1、16.0、11.1、21.9、26.9,上层林冠盖度与薇甘菊盖度的关系是斜率相同(-0.24)、截距不同的平行线(图 1);图1中粗线为总体趋势线,其截距是 6个林分的加权平均;林分A所处生境1的薇甘菊繁殖体压力特别大,混合效应模型将这种林分不同造成的差别体现到截距上,避免因林分不同影响到斜率的估算。

表3 以薇甘菊盖度为响应变量、林分类型为随机截距、单个因子为解释变量的混合效应模型分析结果Table 3 Mixed effect modeling with Mikania micrantha cover as response variable, stand factor as random intercept and single environment as explanatory variable

图1 以薇甘菊盖度为响应变量、林分类型为随机截距、上层林冠盖度为解释变量的混合效应模型Figure 1 Mixed effect modeling with Mikania micrantha cover as response variable, stand factor as random intercept and upper canopy cover as explanatory variable
2.4 薇甘菊二元变量的影响因子分析
相关分析表明,薇甘菊开花数量与上层林冠盖度极显著负相关(R= -0.537,α < 0.01)。以薇甘菊有花无花为因变量的二元协方差分析表明,薇甘菊盖度对是否开花有略微显著的正效应(截距B= 0.043,P= 0.065),上层林冠盖度对是否有花有略微显著的负效应(B= -0.04,P= 0.081),其他因子的效应不显著。
3 讨论
3.1 影响薇甘菊盖度的主要环境因子
因生境、林龄、树种等因素的作用交集在一起,难以厘清各环境因子对薇甘菊盖度的效应;林分因子涵盖了生境、林龄和树种的变化信息,以它作为随机截距的单因素混合效应模型分析结果,显示的是林分效应处于均值时薇甘菊盖度与各环境因子的关系,产生适合整个研究区的普适性结果。在林分效应均化的前提下,环境因子与薇甘菊的相关性排序为: 林下植被盖度>上层林冠盖度>朝向>物种丰富度>坡度(表 3),前三因子与薇甘菊盖度的相关性显著。林下植被盖度与薇甘菊盖度正相关,干热朝向与薇甘菊盖度显著负相关。上层林冠盖度是抑制薇甘菊盖度扩大的主要因素,上层林冠盖度每增加10%,薇甘菊盖度减少 2.4%;越是在干热的立地,上层林冠盖度对薇甘菊入侵的阻止效应越明显;因此,薇甘菊疫区造林宜选择能够形成浓密乔木林冠的树种。
本文发现薇甘菊盖度在林下植被盖度高、上层林冠盖度较低、阴湿朝向的立地下较高,这一结果与黄忠良等[8]薇甘菊喜光、好湿的结论基本相符。薇甘菊在郁闭度大、光照少的林分内不易生长[6,23]。但也有观察到薇甘菊盖度与其它植物总盖度、密度显著负相关的报道[24]。
3.2 较大的上层林冠盖度抑制薇甘菊开花和生长
开花结籽是植物生活周期的一个重要阶段,对种群的延续和扩大分布具有重要作用[1]。相关分析和二元协方差分析表明,薇甘菊开花数量与上层林冠盖度极显著负相关(R= -0.537,α < 0.01),上层林冠盖度对薇甘菊开花具略微显著负效应(P= 0.081)。杨期和等[25]在东莞观察到薇甘菊在开阔生境中花数多、花期长,在林荫处花期短、花数少;薇甘菊种子生产需要较充足的光照[11];在较郁闭的群落中,薇甘菊产生较少的花和种子[1];这些都与本文结果大体相符。但也有学者观察到相反情况,如洪岚等[26]在东莞植物园发现开阔地中薇甘菊倾向于克隆繁殖,而林窗生境中倾向于有性生殖。
3.3 源于林龄、生境和造林树种的植被差异表现
本研究的人工林林龄 0.5—3 yr,林龄越大上层林冠盖度越高,新造林的上层林冠盖度接近零。三个生境的薇甘菊繁殖体压力差异较大;在林分A杂交相思的林缘,薇甘菊生长极其茂盛,导致多重比较中林分 A的薇甘菊盖度显著高于林分 B、D、F和C(表2);如果得出杂交相思林薇甘菊入侵程度显著高于同龄桉树纯林的结论,在造林树种选择上将进退两难。上层林冠盖度分箱成低、中、高三个等级,均值比较产生上层林冠盖度中等的样方薇甘菊盖度较高的结果,因生境固有的繁殖体压力不同极易产生误导结论;在生态观测型研究的数据分析中,混合效应模型似乎必不可少,其结果比较科学合理。生境1的杂交相思林和火力楠林因造林树种不同,两个林分的上下两层植被盖度、薇甘菊入侵程度均显著不同,在抵御薇甘菊入侵方面火力楠显著优于杂交相思。
3.4 对薇甘菊疫区造林树种选择的意义
许多研究人员提出利用薇甘菊不耐荫蔽的特性,选择冠幅增长较快的造林树种,营建郁闭林分,达到长期有效控制薇甘菊危害的目的。本研究的三个3.5 yr林分中,桉树林的上层林冠盖度均值显著低于火力楠林和杂交相思林。桉树具有幼态叶、成龄叶两型性,其成龄叶基部偏斜、叶柄扭转、垂直悬挂,因而成龄桉树的树冠很稀疏、很少遮荫[27]。在无薇甘菊侵害的桂南地区,速生桉成龄林的上层林冠盖度一般小于55%。新造半年的桉树幼树是塔形的、地表盖度可能大于 80%,因为幼态叶是平展着生、宽卵形状;当桉树长到2 m高以后,枝条上开始出现中间过渡类型的叶子和成龄叶,不论如何密植都难以提高成年桉树林的上层林冠盖度,成年桉树都可能发生严重的薇甘菊危害。疫区桉树林需要比火力楠和相思林更为频繁地进行薇甘菊防治才能有收获,因此,我们认为薇甘菊疫区不宜种植桉树。
