保护性耕作对西北旱区丛枝菌根真菌及土壤理化性质的影响
2020-10-28王威雁温晓霞李亚君廖允成
杨 华,李 彤,王威雁,温晓霞,李亚君,廖允成
(西北农林科技大学 农学院,陕西杨凌 712100)
土壤耕作对农业生产有着极其关键的影响,对土壤结构、土壤肥力以及土壤生态产生比较显著的影响[1]。保护性耕作与常规耕作方式相比,一方面能够降低劳动力的需求,减少成本投入,另一方面能够使土壤具有稳定的土层结构,优化土壤的理化性质,有效抑制土壤的水分以及营养流失,促进土壤有益微生物的繁殖生长。而常规耕作处理会对土壤的理化性质产生干扰,破坏土壤中微生物的群落结构[2]。研究表明,丛枝菌根(Arbuscular mycorrhizae ,AM)真菌可以与大部分陆地植物的根系构成共生系统[3]。就农作物而言,AM真菌可以促进其根系吸收水分和矿物质营养[4],增强农作物在生物胁迫下的生存能力[5]以及对恶劣环境的抵抗力[6],能够在一定水平提高植物多样性[7]。研究表明,AM真菌易受农作措施的影响,比如耕作措施、施肥条件[8-10]。保护性耕作下土壤的土层结构相对稳定,AM真菌生物量较高,侵染效果明显[11],AM真菌群落丰度提高[12],物种多样性增加[11,13]。土壤pH、有机碳含量、速效磷含量等是影响AM真菌群落结构差异的重要因素[14]。此外,Davison等[15]通过深入研究NS31-AM1引物区段,系统地研究了每一个公开的AM真菌序列,在序列相似性97%的精度下划分出356个AM真菌分子种,以Schüler建立的分类系统为依据,创建了Maarj AM数据库(https://maarjambo-tany.ut.ee/),将孢子种相应区段的DNA序列作为参考,将划分出的分子种归类至属甚至种的水平,称之为虚拟种(Virtual taxa,VT)。目前,高通量测序越来越多地应用到AM真菌研究中[16-17]。
目前,研究主要集中在AM真菌提高作物的养分(尤其是磷肥)吸收、抗逆性等方面,而本试验从保护性耕作对AM真菌影响的角度出发,研究土壤耕作方式对AM真菌的影响以及主要因素。研究长期保护性耕作下农田土壤中AM真菌的生物量、侵染效果及群落结构的差异,对选择合理的耕作方式、充分利用AM真菌资源提供理论依据。试验组采用免耕和深松耕,对照组采用翻耕,结合大田观测和ILLumina Miseq高通量测序技术,探讨长期连续免耕、深松耕和翻耕对土壤中AM真菌生物量、侵染效果和群落结构的影响及其与土壤理化性质的响应关系。
1 材料与方法
1.1 试验地概况
试验地在陕西省杨凌区西北农林科技大学北校区西区,该地区位于渭北旱塬,主要气候为暖温带季风性气候,海拔高度446 m,年日照时长大约为2 150 h,年平均气温为12 ℃~14 ℃,每年约有221 d的无霜期,降雨量与蒸发量分别为621.6和993 mm,试验选用的土壤为塿土。
从2009年开始,试验田采用一年两熟的复种模式,分别种植冬小麦和夏玉米,设置两种保护性耕作:秸秆还田免耕(简称免耕,ZT)和秸秆还田深松耕(简称深松耕,CPT)。同时设置一种常规耕作方式:秸秆还田翻耕(简称翻耕,PT)。每种耕作处理设置3个重复,共划分9个小区,每个小区的面积为375 m2(15 m×25 m)。在玉米播种前,所有处理均施用尿素[w(N)>46%]和磷酸钙[w(P)>16%]750 kg·hm-2。
1.2 样品采集
土壤样品及玉米根系样品采集时间为2018-08-15,此时玉米正处于吐丝期。采用五点取样法,在无根区域使用土钻采取2~30 cm的土壤,混合均匀后过1 mm筛,并将过筛后的土壤放入无菌自封袋中,作为该耕作处理的一个样本。所有土壤样品在低温冰盒内带回实验室,一部分保存于-80 ℃冰箱,用于土壤微生物总DNA提取;一部分放在自然通风的环境下风干,用于土壤理化性质、孢子数、菌丝密度的测定。从2~30 cm土层取玉米新生根样10 g左右,用清水清洗,放置在FAA固定液中,用于测定侵染率。每个处理3个重复,土壤和根系各采集9个样品。
1.3 土壤理化指标的测定
土壤理化性质的测定方法如下:利用激光粒度仪测定土壤粉粒、黏粒及砂粒所占比重;利用电位法进行土壤酸碱度的测量(水土体积比例为 5∶1);利用凯氏定氮法测定土壤全氮含量[2,18];土壤中硝以及铵态氮含量利用2 mol·L-1KCL浸提,使用全自动间断分析仪测定[19];利用重铬酸钾外加热法测定土壤中有机碳含量;利用NaHCO3浸提-钼锑抗比色法测定土壤速效磷含量[20]。
1.4 AM真菌生物量和侵染效果的测定
取新鲜玉米根系,剪成长度为1 cm的根段放入根盒中,利用醋酸墨水进行根段的染色[23],染色过程结束后,使用清水进行根段的清洗,在显微镜下观察。根据菌根侵染和丛枝丰度分级标准将等级参数输入到MYCOCALC软件中,利用该软件进行菌根侵染频率,整个根系的菌根侵染强度和根系丛枝丰度的计算[22]。
1.5 土壤总DNA提取与PCR扩增
按照E.Z.N.A.soil试剂盒操作规范抽提土壤DNA,使用NanoDrop2000测定DNA质量浓度与纯度。所有样品的AM真菌DNA采用巢式PCR扩增。第一对引物:AML1F(5′-ATCA ACTTTCGATGGTAGGATAGA-3′)和AML 2R(5′-GAACCCAAACACTTTGGTTTCC-3′),第二对引物:AMV4-5NF(5′-AAGCTCGTAGTTGAATTTCG-3′)和AMDGR(5′-CCCAACTATCCCTATTAATCAT-3′)[24]。PCR扩增程序如下:98 ℃5 min;98 ℃45 s,58 ℃50 s,72 ℃ 45 s,循环30次;72 ℃延伸10 min[25]。
1.6 Illumina Miseq测序
使用20 g·L-1琼脂糖凝胶回收PCR产物,利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City,CA,USA)纯化,Tris-HCl洗脱,琼脂糖凝胶电泳检测。使用QuantiFluorTM-ST进行定量检测。根据Illumina MiSeq平台(Illumina,SanDiego,USA)标准操作规程纯化扩增片段,构建PE 2*300文库。测序在Miseq PE300平台进行(上海美吉生物医药科技有限公司)。
1.7 数据处理与分析
使用Excel 2007和SPSS 21进行数据整理、统计分析,使用Excel 2007进行绘图。利用Person相关系数评价土壤理化性质与侵染效果以及AM真菌生物量之间所存在的内在关联。高通量测序原始序列通过Trimmomatic软件质控,使用FLASH软件拼接。使用UPARSE 软件(version 7.1)对所有序列进行OTU聚类(97%相似度),并将嵌合体及单序列去除。所得序列利用RDP classifier进行物种分类注释,与Silva数据库(SSU123)比对,比对阈值为70%。按照最小样本序列数对测序所得数据抽平处理后,进行AM真菌群落结构分析,包括Alpha多样性、物种组成分析、基于Bray-Curtis算法的Beta多样性分析以及利用单因素方差分析(One-way ANOVA)进行物种差异分析。同时采用冗余分析(RDA分析)进行土壤理化性质与AM真菌群落结构的相关性分析。数据以“平均值±标准误”表示。
2 结果与分析
2.1 不同耕作处理对土壤理化性质的影响
由表 1 可知,不同耕作方式改变土壤的理化性质。翻耕的土壤中黏粒含量显著高于深松耕(6.70%)。深松耕和免耕的土壤中粉粒含量显著高于翻耕(2.34%和1.75%),深松耕和免耕的土壤中粉粒含量无显著差异。深松耕的土壤中砂粒含量显著高于免耕(56.98%)和翻耕(73.46%),免耕与翻耕的土壤中砂粒含量无显著差异。翻耕的土壤pH显著高于深松耕(1.91%)和免耕(2.30%),深松耕和免耕的土壤pH无显著差异。深松耕与土壤中有机碳含量高于免耕(8.57%),免耕高于翻耕(28.36%),三者差异显著。深松耕的土壤中全氮含量高于免耕(14.97%),免耕高于翻耕(15.75%),三者差异显著。三种耕作处理土壤中的硝、铵态氮含量无显著差异。翻耕的土壤中速效磷含量高于深松耕(15.79%),深松耕高于免耕(19.16%),三者差异显著。
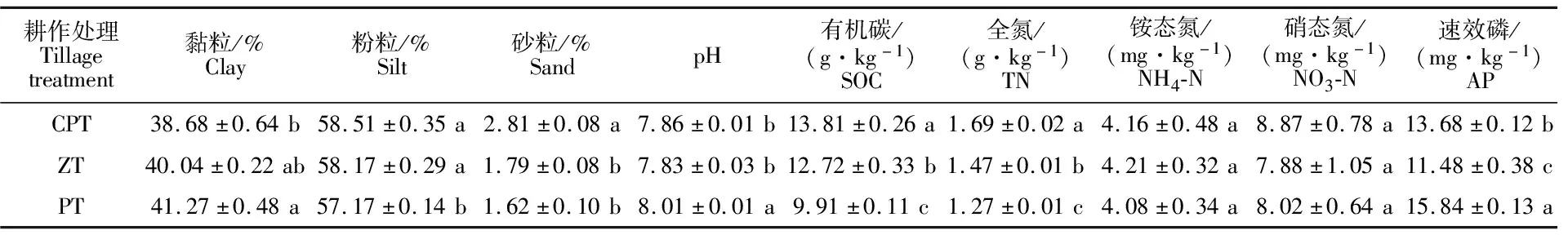
表1 不同耕作处理下土壤的理化性质
2.2 不同耕作处理对AM真菌生物量的影响
如图1-A所示,20 g 土壤中,免耕处理的孢子数显著高于深松耕(24.82%),深松耕土壤中孢子数显著高于翻耕(82.89%)。由图1-B可知,免耕和深松耕土壤中菌丝密度显著高于翻耕,分别高57.04%和43.17%,免耕与深松耕无显著性差异。

不同字母表示差异显著(P<0.05);下同
2.3 不同耕作处理对AM真菌侵染效果的影响
如图2所示,免耕和深松耕条件下AM真菌对玉米根系的侵染率显著高于深松耕(2.17%和35.57%);免耕条件下AM真菌对玉米根系的侵染密度显著高于深松耕(17.19%),深松耕下侵染密度显著高于翻耕(42.33%);免耕条件下AM真菌在玉米根系中的丛枝丰度显著高于深松耕(28.36%),深松耕下AM真菌在玉米根系中的丛枝丰度显著高于翻耕(50.09%)。
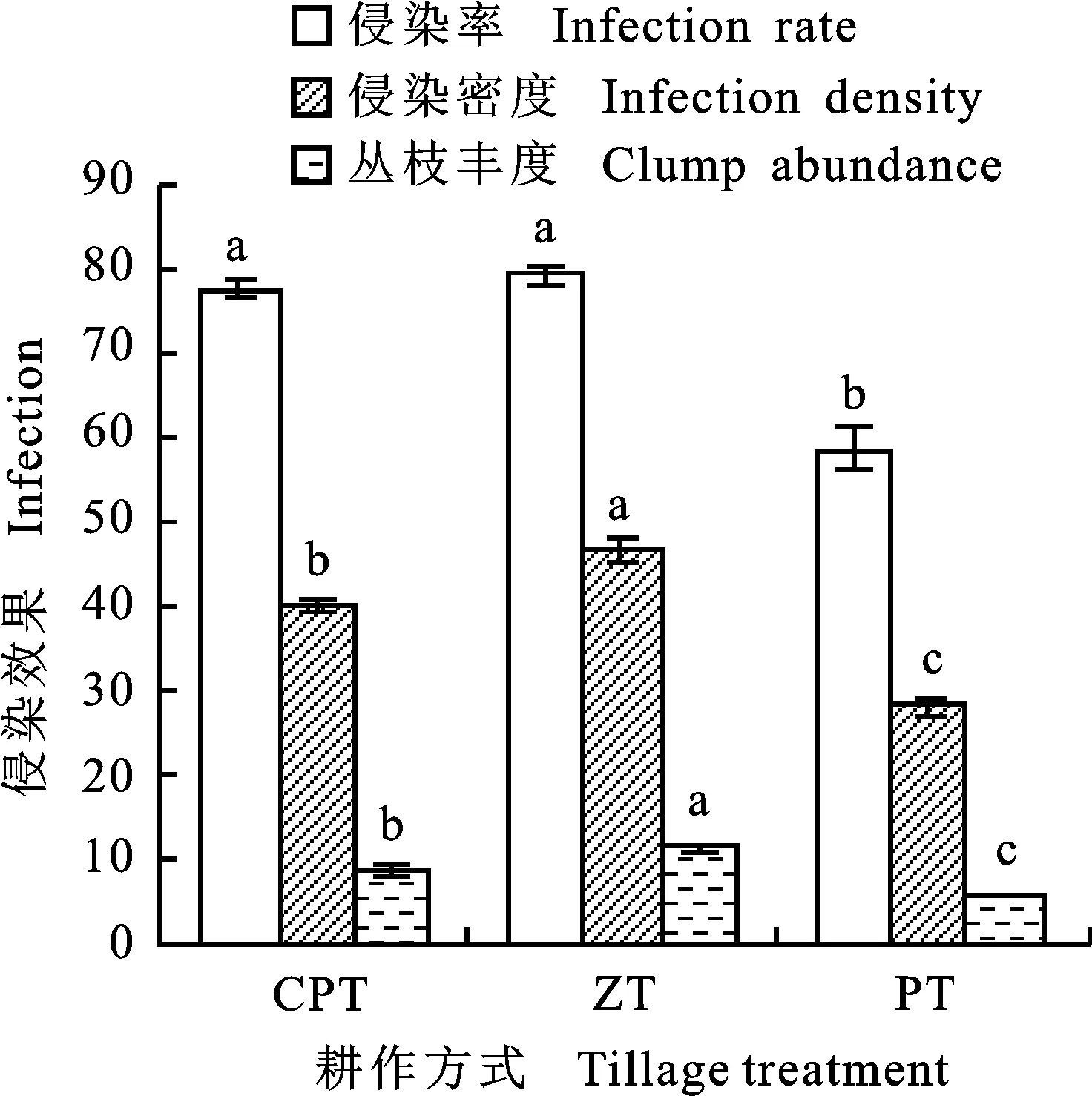
图2 不同耕作处理下AM真菌对玉米根系侵染效果的差异
2.4 土壤理化性质与AM真菌生物量、侵染效果的相关性
如表2所示,土壤样品的粉粒含量和与侵染密度以及侵染率之间呈正相关,且具有显著性(P<0.05);土壤样品的pH与侵染效果以及AM真菌生物量之间呈负相关系,且具有显著性(P<0.01);土壤样品的有机碳含量与AM真菌的各项指标之间均呈正相关关系,且具有显著性(P<0.05);土壤样品的全氮含量与侵染率及菌丝密度呈正相关,且具有显著性(P<0.05);土壤样品的速效磷含量与AM真菌的所有指标都存在着负相关关系,且具有显著性(P<0.01)。
《庄子·天道》:……水静犹明,而况精神!……夫虚静恬淡,寂寞无为者,天地之本,而道德之至,故帝王圣人休焉。休则虚,虚则实,实则备矣。虚则静,静则动,动则得矣。

表2 AM真菌生物量、侵染效果与土壤性质的相关性
2.5 不同耕作处理对AM真菌群落结构的影响
通过高通量测序产生高质量序列共计 495 424条,平均每个测序样本含有序列55 047条,所有序列可以划分为60个OTUs,其中有效序列488 671条,占总序列数的98.67%。分析溶解曲线可知,在相似度为97 %的情况下,各个样品所对应的覆盖率指数(Coverage value)都超过99 %水平,说明当前测序工作的深度满足评价该土壤AM真菌群落结构多样性的要求。
根据Maarj AM数据库(https://maarjambo-tany.ut.ee/)中对AM真菌虚拟种的归类,本试验共鉴定发现AM真菌包含4个属的25个虚拟种(表3)。不同耕作处理下,土壤AM真菌属和虚拟种数量不同。其中在深松耕处理下,土壤中鉴定到3个属的17个虚拟种;免耕处理下,土壤中鉴定到4个属的19个虚拟种;翻耕处理下,土壤中鉴定到2个属的12个虚拟种。

表3 不同耕作处理下AM真菌的属和虚拟种
对不同耕作处理下土壤AM真菌的Alpha多样性指数进行方差分析,结果如表4所示,在OTU水平上,深松耕土壤中AM真菌的Sobs值显著高于翻耕,说明深松耕土壤中实际观测到的OUT数目高于翻耕土壤。深松耕和免耕处理下,土壤中AM真菌的Shannon值显著高于翻耕土壤,说明深松耕、免耕土壤中AM真菌的多样性高于翻耕土壤。深松耕土壤中AM真菌的丰富度指数(Chao1)显著高于翻耕,说明长期深松耕处理下AM真菌的丰富度高于翻耕土壤。

表4 不同耕作处理下AM真菌AlpHa多样性(OTU水平)
2.6 AM真菌群落结构与土壤理化性质的相关性
如图3所示,同一耕作处理的3个样本的群落结构相似,且与其他耕作处理不同。ANOSIM分析表明,不同耕作处理间AM真菌群落的组间差异大于组内差异(R=0.37),且不同耕作处理AM真菌群落结构的差异达到显著水平(P<0.05)。冗余分析结果显示,本试验测定的土壤理化性质共解释81.33%AM真菌群落结构的差异。其中,第一排序轴解释群落结构差异的 46.0%,第二排序轴解释16.1%。蒙特卡洛检验(Monte Carlo检验)结果显示,有机碳(F=4.3,P=0.002,解释量为38.2%)、速效磷(F=2.4,P=0.006,解释量为17.7%)对AM真菌群落结构有显著影响。
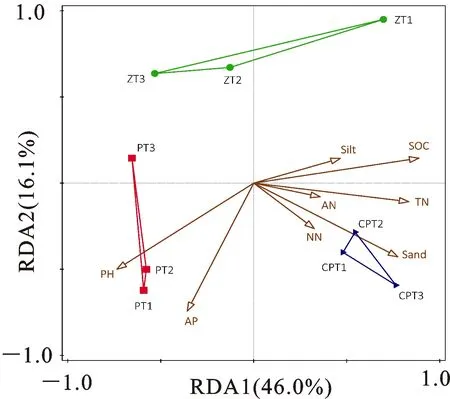
SOC.有机碳;TN.全氮;AN.铵态氮;NN.硝态氮;AP.速 效磷
3 讨 论
3.1 保护性耕作对土壤理化性质的影响
不同的耕作处理对土壤的理化性质产生影响。保护性耕作减少了对土壤的扰动,有助于形成稳定的土层结构。深松耕和免耕的土壤中粉粒含量提高,黏粒含量降低,pH降低。此外,保护性耕作减少了有机质的矿化作用,有助于促进有机质积累[1],与前人的研究结果一致。不同耕作措施对速效磷含量的影响一致存在争议,本试验结果与张贵云等[26]、田慎重等[27]研究结果不同,但与贾凤梅等[28]、李永平等[29]研究结果一致。
3.2 保护性耕作对土壤真菌生物量、侵染及群落结构的影响
农田耕作措施对土壤的扰动对大多数AM真菌来说可能是不利的[30],本研究表明,长期免耕、深松耕处理下,土壤中孢子数量、菌丝密度、侵染率、侵染密度以及根系丛枝丰富度皆高于翻耕。土壤耕作处理可以破坏菌丝网络,直接影响AM真菌的生长、繁殖,降低其侵染能力[31-32]。
本试验只检测到4个属,即球囊霉属(Glomus)、无梗囊霉属(Acaulospora)、多孢囊霉属(Diversispora)以及巨孢囊霉属(Gigaspora)。其中,球囊霉属(Glomus)在土壤中丰度极高,这与前人研究结论一致[19],而其他属丰度很低。球囊霉属(Glomus)的优势说明该属在农田条件下具有较高的生存能力。主要原因在于,相比较于其他AM真菌种类,球囊霉属(Glomus)不仅可以通过孢子繁殖,还可以通过残留的菌丝片段进行繁殖[33],而巨孢囊霉属(Gigaspora)只能通过孢子散布或通过完整的菌丝来侵染繁殖[34]。此外,Giovannetti等[35]的研究已经表明,Glomus属很容易在菌丝之间形成连接,因此在农田土壤受到机械破坏后可以较快地重建菌丝网络,恢复自身的生长与繁殖。这也解释了本试验中土壤耕作(免耕、深松耕、翻耕)对Glomus属的优势地位未产生影响,但是,免耕土壤中保存了更多AM真菌的种类,而剧烈的土壤扰动使翻耕土壤中有些AM真菌种类在本次测序结果中检测不到。
尽管在属水平3种耕作处理土壤中AM真菌的群落结构丰度无明显差异,但是在分子种及OTU水平仍可以发现不同耕作处理对AM真菌群落结构的多样性产生一定影响,对于保护存在于土壤里的AM真菌而言,深松耕及免耕的方式作用明显。
3.3 不同耕作处理下,AM真菌生物量、侵染及群落结构与土壤理化因子的关系
土壤pH与AM真菌生物量及侵染效果均呈显著负相关。以往的研究结果显示,AM真菌产孢在一定程度受到土壤pH的影响,进而对AM真菌多样性产生作用[36-37]。在pH 7.8~10.5随着pH升高,AM真菌孢子数降低,不同种类对其抗性有所差异[38]。
有机碳含量与AM真菌生物量及侵染效果呈显著正相关。冗余分析结果表明,土壤中AM真菌的群落结构也受到有机碳的显著影响。有机质可以为AM真菌提供生长环境,提高其侵染能力,有机碳含量增加可以促进AM真菌分枝,AM真菌菌丝倾向在土壤有机碳中或者有机碳周围聚集并增殖[39]。此外,如果在土壤中存在比较高的有机质含量,则会促进菌根的发育以及菌丝的生长,然而这种促进作用是有一定限制的[40],在一定区间内,土壤中有机质含量越丰富,AM真菌的种类就越多,但超过阈值后,AM真菌的分布就随着有机质含量的增加而减少[41]。
速效磷含量与AM真菌生物量及侵染效果呈显著负相关,冗余分析结果表明,土壤中AM真菌群落结构受到速效磷含量的显著影响,与前人研究结果一致[42]。在营养充足的土地中增施P肥则减少孢子数量、菌丝分枝和菌丝总长度[43-44]。如果土壤里的磷含量比较高,植物和AM真菌的共生关系将会出现利益分配的改变,植物对AM真菌提供的物质与能量减少[22],将会在一定程度上抑制其泡囊发育[43]。在长时间的植物选择之后,部分物种因无法获得充足能量供给而逐渐被自然界淘汰,所以在翻耕处理的土壤里,速效磷相对比较高的含量将会导致AM真菌多样性的降低。
总之,常规耕作方式下,定期翻耕引起土壤理化性质改变,提高土壤pH,使土壤中有机碳的含量降低,而速效磷的含量增加,改变土壤中AM真菌群落结构组成,降低AM真菌在作物生长过程中的贡献。相反,保护性耕作如免耕、深松耕为AM真菌的生长提供稳定的生存环境,在一定程度上保护AM真菌的多样性。
本研究虽以10 a长期定位试验为基础,但取样只有1 a的吐丝期,没有覆盖全部生育期。在以后的研究中将增加关键生育期并增设取样年份,以便获得更全面的研究结论和动态结果。
