白菜型冬油菜 ELIP1基因的克隆与低温表达分析
2020-10-28何辉立孙柏林刘丽君武军艳孙万仓金娇娇李学才
何辉立,方 彦,马 骊,孙柏林,刘丽君,武军艳,董 云,4,孙万仓,金娇娇,李学才
(1.甘肃省干旱生境作物学重点实验室, 兰州 730070; 2.甘肃省油菜工程技术研究中心, 兰州 730070; 3.河北中旭检验检测技术有限公司, 石家庄 050200;4.甘肃省农业科学院作物研究所,兰州 730070)
植物早期光诱导蛋白(early light-induced protein, ELIPs)是核基因编码的叶绿体类囊体膜蛋白[1],属于叶绿素结合蛋白超家族(chlorophyll a/b binding superfamily, CABs)。其最早发现于黄化豌豆幼苗去黄化试验中[2],具有光保护作用[3]。ELIP蛋白也是一种逆境胁迫响应蛋白,植物在受到高盐[4]、干旱和低温等多种非生物胁迫时体内ELIP被诱导且表达量提高[5]。低温胁迫是ELIP基因表达的主要诱导因素之一[3]。低温驯化过程中,豌豆(PisumsativumLinn)[6]、大麦(Hordeumvulgare)[7]、拟南芥[8](Arabidopsisthaliana)和盐生杜氏藻(Dunaliellasalina)[9]等多种物种ELIP基因被诱导表达,对提高植物抗寒性十分重要。Shimosaka等[10]研究发现,小麦中ELIP在低温胁迫过程中被大量诱导产生,具有增强植物抵御抗寒的能力。在冷驯化中诱导表达ELIP基因有助于紫花苜蓿falcata抗寒性的提高[11]。
白菜型冬油菜是北方旱寒区一种重要的越冬经济作物,可在北方不同生态区正常越冬[12],蕴藏着丰富的抗寒基因,是研究植物抗寒性及获得抗寒基因的重要材料。甘肃省干旱生境作物学重点实验室前期在用转录组学分析方法筛选白菜型冬油菜的冷诱导差异基因时,从数据库筛选和鉴定出参与低温诱导途径的ELIP1基因,为进一步研究ELIP1基因在白菜型冬油菜低温胁迫中的功能,本试验克隆8个不同抗寒性冬油菜品种叶片中的ELIP1基因,运用生物信息学方法预测该基因的结构和功能,并利用实时荧光定量PCR技术研究不同抗寒性冬油菜品种在低温胁迫下的表达模式,研究结果有助于理解ELIP1基因在冬油菜抗寒性中的作用,并对后期开展的抗寒育种有一定参考意义。
1 材料与方法
1.1 材 料
选取8份白菜型冬油菜为试验材料,来源及特性见表1。
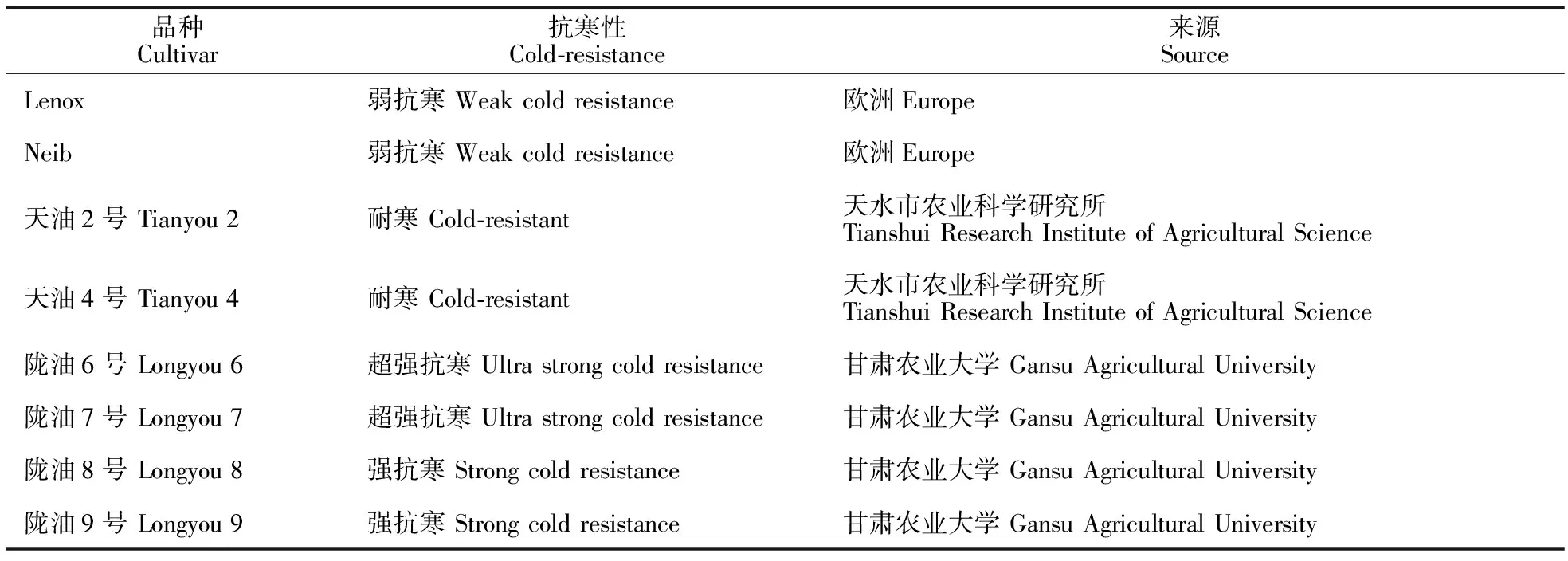
表1 品种名称与来源
1.2 试验设计
采用盆栽试验,将籽粒饱满的各品种油菜种子播种至装有育苗基质的营养钵中(9 cm×7.5 cm),每钵4株,放置于甘肃农业大学校园温室大棚中,培养至五叶期。选取健壮、长势一致的不同油菜幼苗在25 ℃(CK,光照14 h,黑暗10 h)和 4 ℃(光照14 h,黑暗10 h)各处理2 d,取第5片真叶用锡箔纸包裹,液氮速冻后放于-70 ℃超低温冰箱保存,备用。
1.3 方 法
1.3.1 RNA提取及反转录 参照天根试剂盒DP419说明书提取各样品总RNA,电泳检测后,按TaKaRa cDNA第一链合成试剂盒(RR036A)说明书进行反转录,得到单链cDNA,超微量紫外分光光度计检测其浓度后置于-20 ℃冰箱保存,备用。
1.3.2 引物设计与ELIP1基因克隆 以实验室前期转录组测序结果为依据,利用Primer Premier 5.0 软件设计cDNA扩增引物,ELIP1-F:5′-ATGGCAATGGCGTCGTTTAACATG-3′;ELIP1-R:5′-TCAAACGAGAGTCCCACCCTTGA-3′,以4 ℃低温处理后各品种叶片提取RNA反转录的cDNA为模板进行PCR扩增,扩增条件:94 ℃ 5 min;94 ℃30 s,60 ℃30 s,72 ℃1 min,35个循环;72 ℃10 min;4 ℃保存。用10 g/L琼脂糖凝胶电泳检测PCR扩增产物片段大小,普通琼脂糖凝胶DNA纯化回收试剂盒(离心柱型)回收目的条带,将其连接到pMD-19T载体上,转化至大肠杆菌DH5α感受态细胞中,在含有Amp、X-gal和IPTG的LB固体培养基上,过夜培养12 h后进行蓝白斑筛选,挑取白斑至含有Amp的LB液体培养基中摇菌。菌落PCR验证的阳性克隆,随机挑选3个送至上海生工生物工程股份有限公司进行克隆结果测序。
1.3.3 ELIP1预测蛋白的生物信息学分析 蛋白质基本理化性质分析采用Protparam;采用Protscale分析氨基酸疏水性。跨膜结构预测采用TMpred Server软件。利用SignalP-5.0 Server预测蛋白信号肽。利用SOPMA工具预测蛋白质二级结构[13]。NCBI CD-search预测ELIP1蛋白保守结构域。用DNAMAN软件比较8个白菜型冬油菜品种中ELIP1基因核苷酸序列的同源性。
1.4 白菜型冬油菜 ELIP1基因的荧光定量表达分析
根据获得的ELIP1基因的CDS序列,设计定量表达引物,ELIP1-F:5′-GAGATGTATGGCTGAG GGAGAA-3′和ELIP1-R:5′-GAGGTGGCGACGATGACTT-3′;内参基因β-actin引物,F:5′-TGTGCCAATCTACGAGGGTTT-3′;R:5′-TTTCCCGCTCTGCTGTTGT-3′,以浓度一致的8个冬油菜品种常温及低温处理后叶片cDNA为模板,进行实时荧光定量分析,PCR扩增条件同上。采用2-ΔΔCt方法进行基因差异表达分析。低温差异表达分析以常温对照(CK)的表达量为基准。3次重复。
2 结果与分析
2.1 ELIP1基因克隆
通过‘陇油7号’的转录组数据分析,发现一条ELIP1基因序列,该基因具有完整编码区的转录本,长度为550 bp。本研究以低温下8个白菜型冬油菜叶片cDNA和特异性引物为模板,用RT-PCR方法扩增,得到一条550 bp左右(图1)的扩增产物,与数据库中ELIP1基因长度吻合。纯化回收目的条带,连接T载体并转化后进行菌液PCR验证挑选阳性克隆(图2),得到的目的片段大小在550 bp左右,说明克隆的ELIP1已插入载体中。将阳性克隆送上海生工生物工程股份有限公司测序,得到长度为588 bp的编码区 序列。
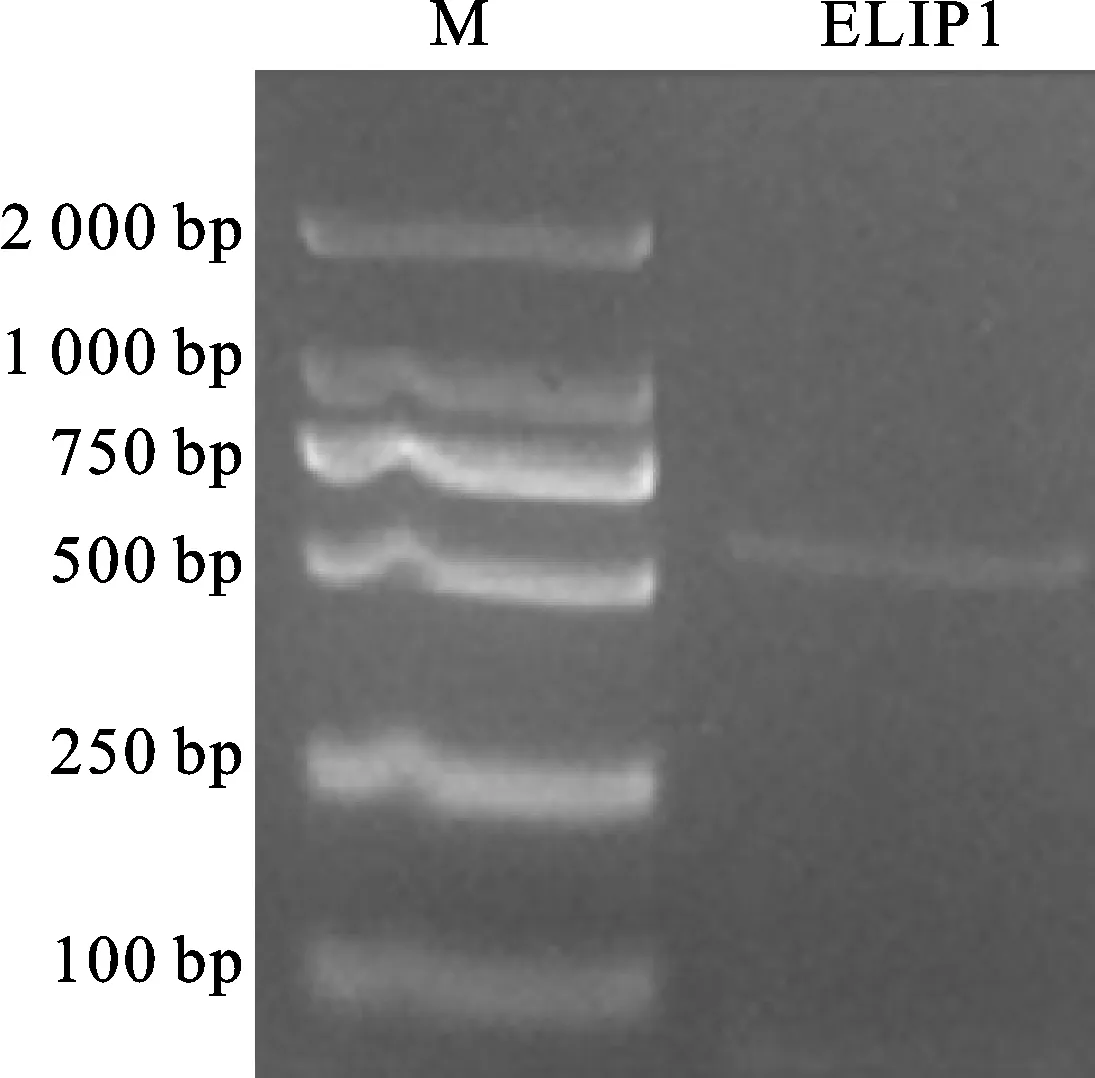
图1 白菜型油菜 ELIP1基因的RT-PCR扩增结果

图2 白菜型油菜 ELIP1基因菌液PCR鉴定结果
2.2 白菜型冬油菜 ELIP1基因编码蛋白质的特性分析
对‘陇油7号’ELIP1基因的碱基序列进行开放阅读框分析,结果显示ELIP1基因CDS区含有一个长度为588 bp 的完整开放阅读框(ORF),编码含195个氨基酸的蛋白质(图3)。蛋白质理化性质分析表明,白菜型冬油菜‘陇油7号’ELIP1蛋白由20种氨基酸组成,其中Ala占11.3%,Leu占10.3%;含有13个酸性氨基酸(D、E),18个碱性氨基酸(K、R),76个疏水氨基酸(A、I、L、F、W、V),47个极性氨基酸(N、C、Q、S、T、Y)及36个带电荷的氨基酸(R、K、H、Y、C、D、E);预测等电点为9.24;相对分子质量为 20.472 8 ku;预测不稳定指数为39.35,是一个稳定蛋白(稳定系数<40时稳定);总平均亲水指数为 0.169,为疏水性蛋白(图4)。

图4 ELIP1蛋白的疏水性预测分析
蛋白信号肽预测分析表明白菜型冬油菜ELIP1蛋白信号肽序列没有断裂(图5)。‘陇油7号’ELIP1蛋白的二级结构(图6)由α-螺旋(alpha hELIP1x,43.08%),延伸链(extended strand,11.79%),β-转角(beta turn,5.13%)和无规则卷曲(random coil,40%)组成。

图5 预测白菜型冬油菜ELIP1蛋白信号肽

蓝色.α螺旋;紫色.无规则卷曲;红色.延伸链;绿色.β转角。横坐标数值表示氨基酸序列位置
利用NCBI CD-search预测ELIP1蛋白保守结构域(图7),预测结果表明ELIP1含有叶绿素A-B结合蛋白家族特有的保守结构域,该保守结构区域是由93(96~188)个氨基酸组成,说明ELIP1是一种参与光合作用的功能蛋白。
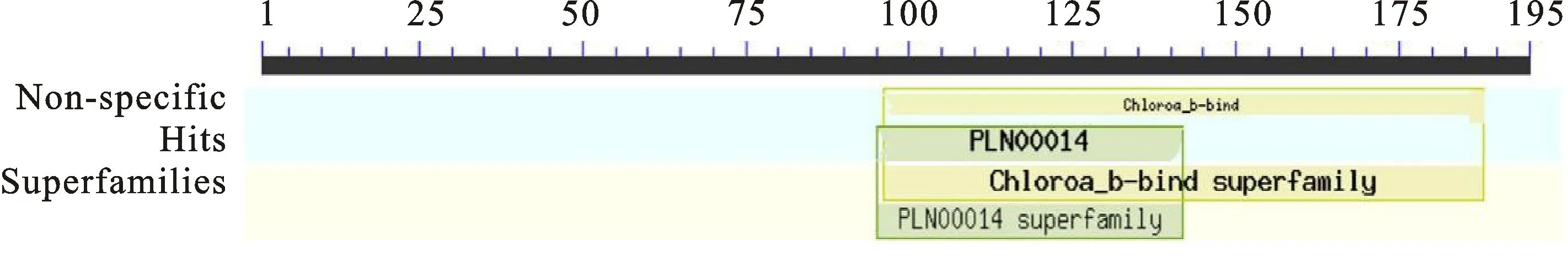
图7 白菜型冬油菜ELIP1蛋白的保守结构域分析
2.3 各品种白菜型冬油菜 ELIP1基因核苷酸序列对比
利用DNAMAN软件对不同白菜型冬油菜品种ELIP1基因进行比较(图8),8个品种ELIP1基因同源性为99.06%。与参考序列白菜型油菜(XM_009137582)对比,ELIP1基因在8个冬油菜品种中共有21处(图8)碱基发生突变,其中弱抗寒品种‘Lenox’和‘Neib’各发生一处突变,强抗寒品种‘陇油8号’碱基突变最多,出现18处,其他5个品种发生15~17处变异,说明不同白菜型冬油菜品种在进化中ELIP1基因序列存在单核苷酸多态性。
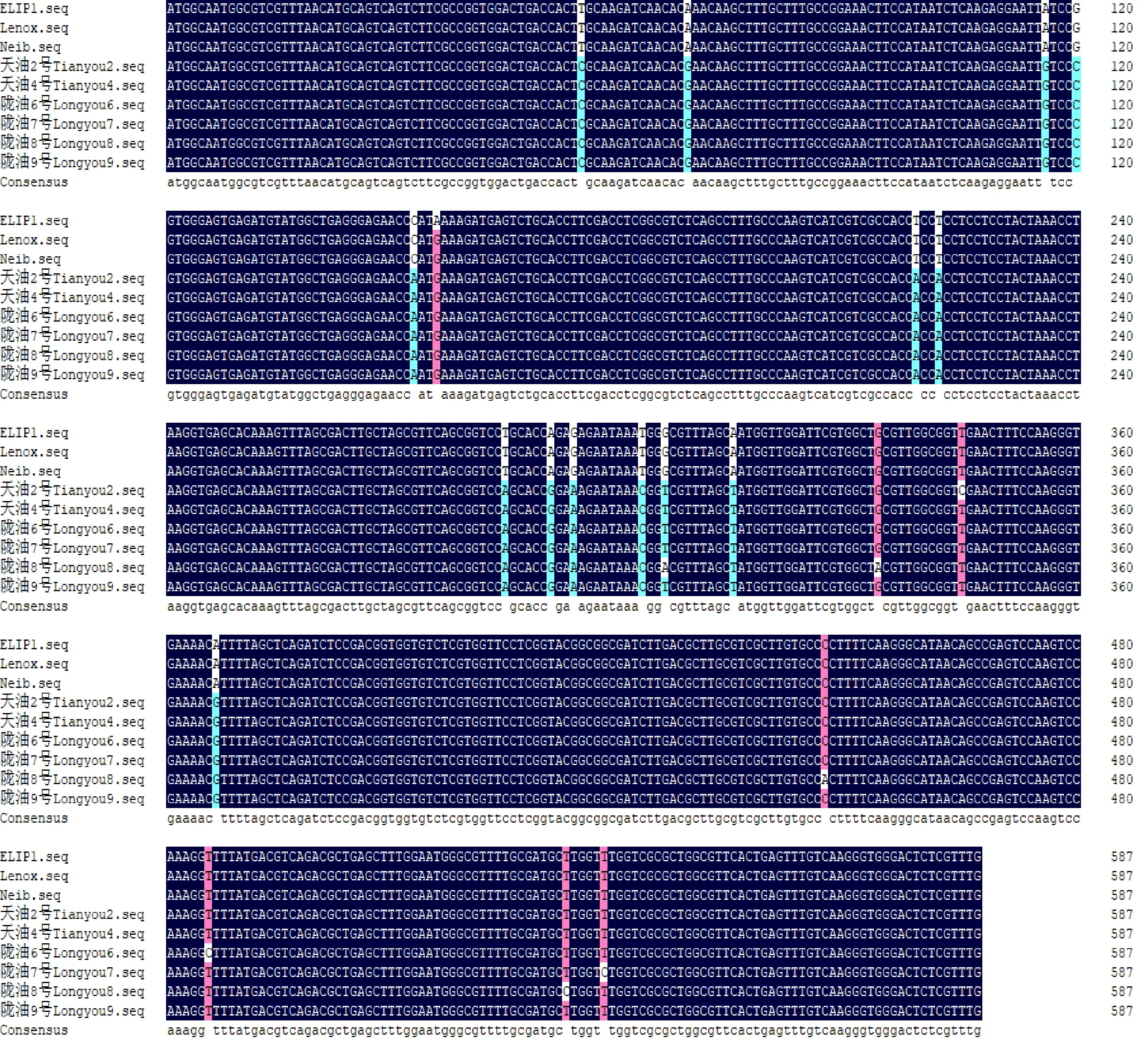
图8 8个白菜型油菜品种 ELIP1序列对比结果
2.4 各白菜型冬油菜品种ELIP1蛋白系统进化树构建与分析
利用MEGA 7.0中邻近法原则对8个白菜型冬油菜ELIP1构建了进化树(图9),结果显示国外品种‘Lenox’和‘Neib’聚为一类;其他6个国内品种聚为一大类,表明国外和国内品种其亲缘关系较远。
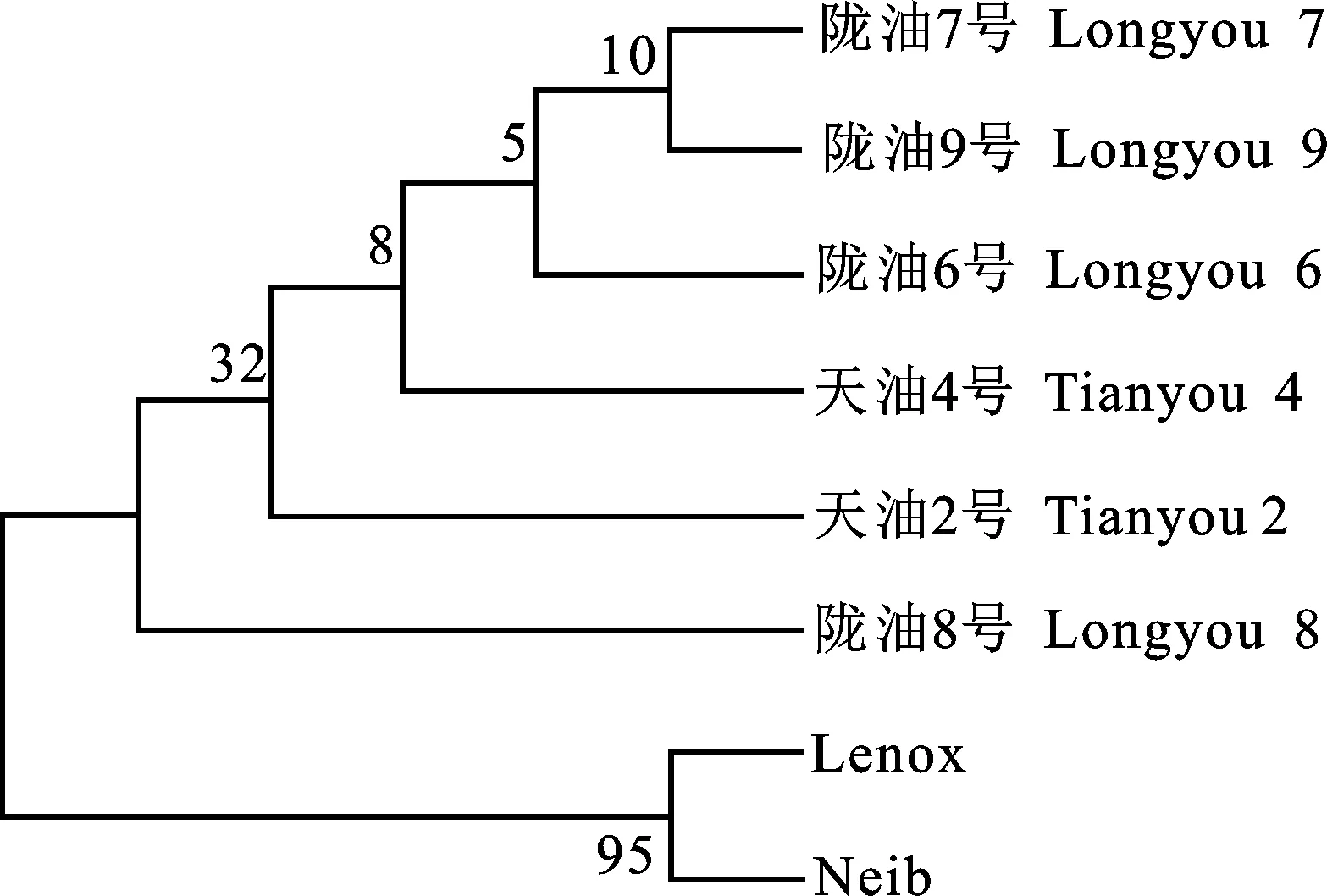
图9 ELIP1蛋白序列系统进化树分析
2.5 低温胁迫对不同冬油菜品种ELIP1表达的影响
通过比较不同冬油菜品种在常温和低温处理后ELIP1 表达水平(图10),发现4℃时,超强抗寒品种‘陇油6号’和‘陇油7号’的ELIP1基因表达量高于25 ℃,分别上调了47.99%和 40.46%,其他品种低温下的表达量均低于常温,且抗寒性越弱,低温下表达量越低。其中抗寒品种‘陇油8号’和‘陇油9号’较25 ℃分别下调了8.88%和5.40%,耐寒品种‘天油2号’和‘天油4号’较25 ℃分别下调了49.14%和40.96%,弱抗寒品种‘Lenox’和‘Neib’在4 ℃时ELIP1基因下调了99.51%和98.28%,表达量很少。

不同小写字母表示不同品种 ELIP1表达量在不同温度处理间差异显著(P<0.05)。
3 结论与讨论
本试验从白菜型冬油菜叶片克隆获得1条含有完整ORF的ELIP1基因,其氨基酸序列与转录组数据库中ELIP1基因序列吻合率为 99.06%,生物信息学分析表明,该基因编码195个氨基酸,等电点为9.24;相对分子质量为 20.472 8 ku;编码主要由a-螺旋和无规则卷曲组成的疏水性稳定蛋白。8个油菜品种核苷酸序列对比分析发现,ELIP1基因存在21处突变,其中弱抗寒品种发生突变位置少于抗寒品种;对8个白菜型冬油菜ELIP1构建进化树进行分析,结果显示两个国外引进弱抗寒品种‘Lenox’和‘Neib’聚为一类,‘天油2号’‘天油4号’‘陇油6号’‘陇油7号’‘陇油8号’和‘陇油9号’聚为一大类,表明国外和国内品种其亲缘关系较远。这可能是弱抗寒品种与抗寒品种存在抗寒性差异的主要影响因素。
目前,已在许多物种中克隆出了ELIP基因[14-15],该基因在受到逆境胁迫时表达量升高,对抵御不同逆境起着重要作用[3]。Montané等[7]研究发现,当温度在5~25 ℃时,随着温度的下降和胁迫时间延长,ELIP基因被诱导程度越高,表达显著上升。Wei等[16]、Peng等[14]、何飞等[17]、Wang等[18]研究发现,低温驯化过程中,植物ELIP基因表达量上调,对提高植物抗寒性十分重要。本研究对8个不同冬油菜品种叶片中ELIP1基因的表达分析显示,4 ℃时,抗寒性越强的品种ELIP1基因表达量越高,表明ELIP1基因可能在冬油菜响应低温胁迫提高自身抗寒性中发挥重要作用。
