不同含油量油菜品种的油脂累积通路上基因的差异表达
2020-10-22于丽欣郑树东柏云会杨加虎陈晶亮杨进成林良斌
于丽欣 郑树东 孙 超 柏云会 杨加虎 陈晶亮 汪 骞 杨进成 林良斌
(云南农业大学农学与生物技术学院1,昆明 650201) (玉龙县农业技术推广中心2,丽江 674100) (云南省农业科学院园艺作物研究所3 ,昆明 650201) (玉溪市农业科学研究院4,玉溪 653100)
菜籽油是一种营养保健的食用油,富含人体必需不饱和脂肪酸,特别是油酸达60%以上,越来越受到人们的青睐。但我国食用油仍有很大的缺口,而油菜(BrassicanapusL)是我国主要油料作物,发展油菜生产是保障我国食用油安全的有效途径。提高菜籽含油量是当前油菜育种和栽培的主要目标之一。油菜种子中的油脂累积是一个十分复杂的生物学过程,涉及到光合作用、糖转运、糖酵解、脂肪酸合成、油脂合成、油体形成等。例如光系统II BY蛋白(photosystem II BY,PSBY)和光系统II反应中心PSB28蛋白(photosystem II reaction center PSB28 protein,PSB28)参与了光合作用,对光系统II中间体的RC47分别起组装保护和激活作用[1,2];蔗糖转运蛋白1(sucrose transporters 1,SUC1)和糖转运蛋白12(sugertransportprotein 12,STP12)参与了糖转运过程,将光合产物从“源”器官运输至种子等“库”器官中[3-4];丙酮酸脱氢酶E1α(pyruvate dehydrogenase E1 α,PDH-E1α)和烯醇化酶1(enolase 1,ENO1)参与了糖酵解过程,PDH-E1α是丙酮酸脱氢酶复合物(pyruvate dehydrogenase complex, PDHC)的重要部分,PDHC进一步将丙酮酸脱氢脱羧,产生脂肪酸合成的原料乙酰辅酶A[5],而ENO1为质体内的合成代谢过程提供烯醇式丙酮酸PEP,经由丙酮酸激酶(pyruvate kinase)的催化,进而生成ATP以及丙酮酸[6];以上这些与含油量的关系研究均鲜见报道。乙酰辅酶A羧化酶1(acetyl CoAcarboxylase 1,ACC1)参与了脂肪酸合成,它将乙酰辅酶A羧化丙二酸单酰辅酶A,是一个脂肪酸合成过程第一个限速酶,在油菜中过表达拟南芥ACC1基因,使T2代种子含油量增加约5%[7]。提高或抑制参与这些过程的酶或蛋白编码基因的表达水平,将会影响油菜种子的含油量,这已被许多基因工程研究的结果所证实[8-16]。一些转录因子调控这些过程中相关基因的表达,从而调控植物种子的含油量[17-21]。一些基因表达和酶活性的研究结果也说明了这一点,如:甘油-3-磷酸脱氢酶基因BnGPDHc2-1、BnGPDHc2-2在高含油自交系30DAF的种子中的表达量显著高于低油含量自交系的[22];BnACC1基因在油菜种子成熟期的相对表达量与油脂积累呈现显著正相关[23];但BnDGAT1、BnPDAT1的qPCR分析说明其表达量与菜籽含油量并不呈现出一致的正相关,这可能与其取材的环境因素不一致有关;在30DAF后的油菜种子中G6PDH、ATP-柠檬酸裂解酶、ACCase、PPase、DGAT的酶活性与菜籽含油量呈显著或极显著正相关[24-27]。对油脂累积通路上的基因表达、酶活性与菜籽含油量相关性的研究仅涉及到几个基因和酶。据不完全统计油脂累积通路上有几百个基因参与,可能不同基因型材料的种子含油量分别是不同的基因在起关键作用。为此,我们选取转录组分析差异表达显著(未发表)的PSB28、PSBY、SUC1、STP12、PDH-E1α、ENO1、ACC1等7个基因进行qPCR及其蛋白或酶活性分析,探究它们在不同含油量油菜品种的角果皮或种子中的表达水平与菜籽含油量的关系,为揭示油菜含油量的分子机制及油菜高含油量育种与栽培提供参考。
1 材料与方法
1.1 材料
植物材料为3个不同含油量的甘蓝型油菜品种:ZY511,湘油15号,湘774,由云南农业大学油菜研究室提供。其中ZY511为浙江农科院选育的常规油菜品种,其原种含油量为50.21%;湘油15号和湘774为湖南农业大学选育的常规油菜品种,其原种含油量分别为42%和37.32%。
1.2 方法
1.2.1 油菜种植与取样
2018年10月,将3种油菜材料种植于云南农业大学后山农场同一块实验地,统一进行油菜常规栽培管理。油菜开花时,对健康、长势一致的油菜植株的主花序上花蕾挂牌并标注日期。一定时间后同时从3个油菜品种上分别取开花后第23、33、43 d的角果,立即带回实验室。在冰浴条件下将种子和角果皮分离,随即分装,每管0.2 g,液氮速冻后-80 ℃冰箱保存备用。
1.2.2 含油量与主要脂肪酸含量的测定
待成熟时收获同一株的菜籽,晒干后除去杂质,将干净的油菜种子送至河南大学,用NIR 5000型近红外光谱分析仪测定种子的含油量和主要脂肪酸的含量。
1.2.3 RNA样品提取与检测
RNA提取过程参照RNA提取试剂盒说明书,取1 μL RNA样品于紫外分光光度计(Nano Drop ND-2000)检测RNA的纯度和浓度,记录A260/280和A260/A230的吸光值;1%的琼脂糖电泳检测RNA。
1.2.4 实时荧光定量PCR的测定
将转录组分析获得的基因序列在NCBI上进行Blast比对分析,用primer 5.0软件根据基因特异序列设计引物,以便排除同源基因的干扰。以100 ng总RNA为模板,按照BestarqPCR RT Kit说明书配制20 μL逆转录反应体系,合成cDNA第一链,并保存备用。Real time PCR 扩增的反应体系为20 μL(DBI Bestar®SybrGreenqPCRmasterMix),程序为94 ℃ 2 min,94 ℃ 30 s,58 ℃ 30 s,72 ℃ 20 s,40个循环,用Agilent Stratagene荧光定量PCR仪Mx3000P进行荧光定量PCR实验,重复3次。相对表达量的计算公式为2-ΔΔct。
1.2.5 粗蛋白液的制备
从-80 ℃冰箱中取出材料放入预冷的研钵中,加液氮后迅速研磨成粉末,将粉末尽量全部转移入10 mL离心管内;加2 mL 4 ℃预冷的提取缓冲液(100 mmol/L Tris-HCl pH=7.3、5 mmol/L DTT、2 mmol/L EDTA、0.5 mmol/L PMSF、300 mmol/L甘油、0.01%体积Triton X-100),振动2 min充分混匀,4 ℃静置1 h;再加10 μL 0.5 mmol/L PMSF并混匀,4 ℃静置1 h;4 ℃、4 000 r/min离心30 min,将上清液转移至2 mL离心管中,用去离子水定容至2 mL,取200 μL粗蛋白液分装至PCR管中,-80 ℃保存待用。
1.2.6 ELISA测定蛋白或酶活性
将粗蛋白液按公司试剂盒说明书并使用多功能酶标仪(VarioskanLUX)进行标准曲线制作及酶活性和蛋白含量测定,重复3次。每克鲜组织的酶活力及蛋白含量(U/g或ng/g)的计算公式为b/a×2 000÷0.2,其中a为样品孔添加的粗蛋白液量,b为根据样品孔所测得OD450值从标准曲线上获得的粗蛋白液样品的酶活力单位或蛋白量。

表1 不同油菜品种的种子含油量与脂肪酸含量/%
1.3 数据处理方法
采用Excel 2010进行数据整理及作图;采用SPSS统计分析软件进行差异显著性分析。
2 结果与分析
2.1 不同油菜品种的种子含油量及其脂肪酸含量分析
对不同油菜品种的种子含油量及其脂肪酸含量进行近红外光谱分析仪检测,结果(表1)表明供试油菜品种的种子含油量差异显著,最高的是ZY511(43.38%),其次为湘油15号(37.23%),湘774含油量最低(34.75%),但其含油量均低于原种4~7个百分点,这可能与油菜成熟期气温过高有关。供试油菜品种的油酸、亚油酸、饱和脂肪酸的含量在品种间差异显著,但芥酸、亚麻酸的含量在品种间差异不显著。并且油酸含量高的品种亚油酸含量低,油酸含量低的品种亚油酸含量高,这可能与油酸是亚油酸的底物有关。
2.2 种子中油脂累积相关基因的差异表达
为了探究STP12、PDH-E1α、ENO1、ACC1的表达与油菜含油量的关系,检测了不同发育时期油菜种子中其RNA和蛋白水平。结果(图1)表明随着种子发育的进程4个基因表达水平均呈上升趋势,而且43DAF时其RNA和蛋白质水平与含油量呈现出一致的关系,即含油量高的材料中其表达水平也高,含油量低的其表达水平也低,表现为ZY511>湘油15号>湘774,而33DAF或之前的基因表达水平并不与含油量呈完全一致的关系,这表明30DAF~40DAF虽然是菜籽油脂累积的快速时期,但并不是菜籽含油量高低的决定时期,而种子发育的后期才可能是菜籽油脂积累的重要时期,基因表达水平决定着菜籽含油量的高低。但不同基因的表达模式存在一定的差异,STP12、PDH-E1α、ACC1的RNA水平随着种子发育的进程显著性增加,但增幅不到3倍,而ENO1的RNA水平在33DAF前的ZY511种子中增加不显著,但到43DAF时急速增加,3个品种均增幅10倍以上。进一步分析发现23DAF、33DAF基因表达的RNA水平与蛋白水平在品种间呈现出不一致的现象,但43DAF则表现出一致性。如STP12的RNA水平在23DAF时湘油15号与ZY511差异不显著,但其蛋白水平ZY511显著低于湘油15号;33DAF时RNA水平差异显著,即ZY511高于湘油15号,但其蛋白水平差异不显著。PDH-E1α、ACC1的RNA水平在23DAF时3个品种间差异不显著,但其蛋白水平差异显著;33DAF时PDH-E1α的RNA水平在湘774与湘油15号间差异不显著,但蛋白水平湘油15号显著高于湘774;33DAF时ACC1的RNA水平ZY511显著高于湘油15号,但其蛋白水平则反之,即湘油15号显著高于ZY511。ENO1的RNA水平在33DAF时ZY511显著低于湘油15号,但其蛋白水平ZY511显著高于湘油15号。引起这种RNA水平与蛋白水平不一致现象的可能有两个原因:一是不同品种的基因中简并密码子使用不同,同一氨基酸在一个品种的基因中使用偏爱密码子,而在另一个品种的基因中使用限制密码子,这样即使RNA水平相同,但其翻译速度不一样,因此,蛋白水平不同;二是由于qPCR的引物是根据基因特异序列设计的,确保定量检测的特异性。而蛋白检测所用的抗体是植物类或拟南芥蛋白的抗体,酶联免疫反应的特异性就会大大降低,ELISA检测的是一类蛋白,而不是某个基因编码的特异蛋白,如果在油菜种子发育早期有多个同源基因低水平表达,而在菜籽含油量决定期只是油脂累积的同源基因表达,就会导致早期(种子发育)检测的RNA水平与蛋白水平不一致,而晚期(油脂累积)检测的一致。
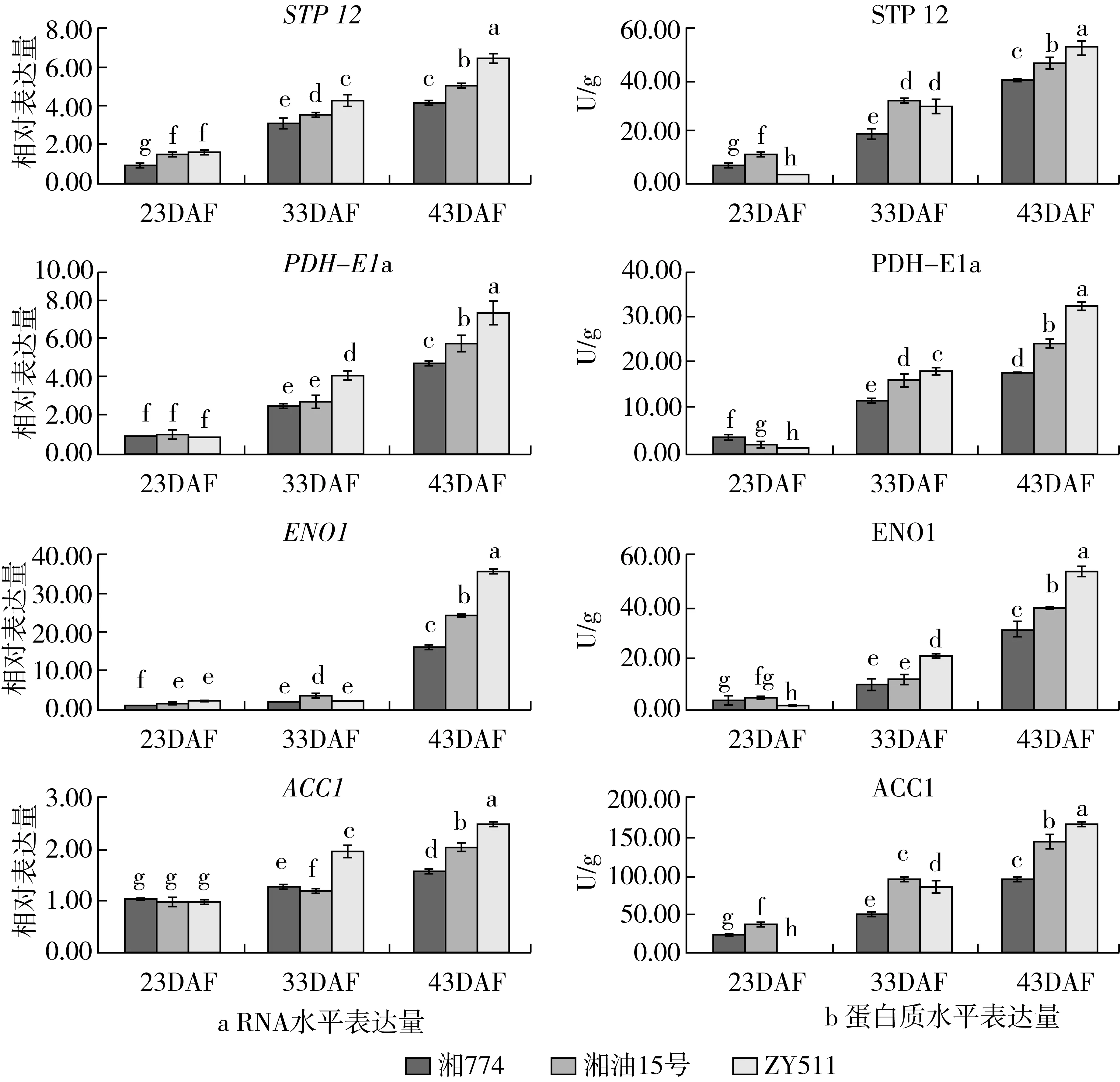
注:图1中的abc均表示为各组多重比较的显著水平为P<0.05。下同。图1 油菜种子中油脂累积相关基因表达
2.3 角果皮中油脂累积相关基因差异表达
为探究油菜角果皮的光合作用与菜籽含油量的关系,本研究选取在不同含油量油菜品种角果皮中差异表达显著的PSBY、PSB28和SUC1等基因进行RNA和蛋白水平定量检测。结果(见图2)表明随着种子发育的进程3个基因表达水平均呈上升趋势,而且43DAF时的RNA和蛋白水平与菜籽含油量均呈一致的关系,即含油量高的材料中其基因表达水平也高,含油量低的材料中其基因表达水平也低,表现为ZY511>湘油15号>湘774,但23DAF、33DAF时的RNA和蛋白水平与菜籽含油量呈现出不一致的关系,这些暗示了43DAF时光合产物和糖转运是专为油脂合成的,而之前特别是23DAF的光合产物和糖转运主要用于角果和种子的生长。3个基因的表达模式存在一定的差异,RNA水平都随着种子发育的进程显著性增加,但蛋白水平PSBY在湘774中呈下降再上升的趋势,其他2个基因与RNA变化趋势一致。此外,PSBY和PSB28的RNA水平增加缓慢,43DAF时最大的增幅也不超过6倍,而SUC1则快速增加,43DAF时3个品种均增幅10倍以上。进一步分析发现角果皮中基因表达的RNA水平与蛋白水平也与种子中基因表达一样,均呈现出品种间不一致的现象。

图2 油菜角果皮中油脂累积相关基因表达
3 讨论
培育高含油量油菜品种是油菜育种的重要目标之一,也是油菜产业增收的重要方式。油脂累积通路上的基因表达水平影响着菜籽的含油量。本研究发现油脂累积通路上的基因表达随着种子发育的进程均呈上升趋势,这可能是因为DNA的甲基化水平随着发育时期的进程其DNA去甲基化逐渐增强,从而导致基因表达水平呈上升趋势。但33DAF或之前的基因表达水平与含油量并不呈完全一致的关系,而43DAF时基因表达水平与油菜种子含油量均呈现一致的关系,说明后期的基因表达水平与菜籽含油量密切相关,是决定油菜含油量的关键时期。这与刘祥含等[27]的研究结果一致,葡萄糖-6-磷酸脱氢酶、乙酰辅酶A羧化酶、磷脂酸磷酸酯酶、二酰甘油酰基转移酶在油菜种子发育过程中随着种子发育进程其酶活性均呈逐渐上升趋势,38DAF时达到峰值,维持一定时间后再逐渐下降。陈玉萍等[24,28]研究表明油菜种子的含油量随种子发育的进程呈上升趋势,从25DAF到30DAF油分积累较慢,30DAF后油分积累较快,种子的含油量迅速上升,40DAF后油分积累减慢,含油量上升较慢。这说明了30DAF~40DAF是油脂快速累积时期,似乎与菜籽的含油量密切相关。本研究结果与其并不一致,可能是虽然30DAF~40DAF是菜籽油脂快速累积期,但并不是含油量的决定期。因此,这一时期的基因表达水平并不与含油量呈一致的关系。40DAF后才是油菜含油量的决定期,油脂累积通路上基因表达水平的高低决定了菜籽含油量的高低。本研究的qPCR检测结果表明随种子发育进程基因表达的RNA水平是逐渐上升的,这与虢慧等[29]研究结果不一致,她们发现在不同含油量油菜品种的种子发育过程中BnACC1、BnPDAT1、BnPDAT1的表达呈波浪式变化趋势,这可能与取材时的环境条件不同有关。在本研究中还发现33DAF或之前的基因表达的RNA水平与蛋白水平不一致,这可能与蛋白检测的ELISA法所选用的抗体特异性有关。由于本研究的ELISA试剂盒的抗体为植物或拟南芥某一蛋白的抗体,而不是油菜某一基因编码蛋白的抗体,这样抗原-抗体反应特异性就会降低,检测到的可能是基因家族的同源基因的编码蛋白,而不是某一基因编码的特异蛋白,从而导致检测结果的RNA水平与蛋白水平不一致。因此,在今后研究中,如要采用ELISA法检测某一特异蛋白,应要通过基因工程的方法制备出这一基因的特异蛋白,然后去免疫动物获得特异抗体。此外,甘蓝型油菜是异源4倍体,几乎在其基因组中每一功能基因都存在多个拷贝。因此,qPCR的引物设计需要格外注意其特异性,否则就会导致检测结果的不准确。
4 结论
本研究利用qPCR和ELISA技术,以3个不同含油量的甘蓝型油菜品种为材料,研究发现油脂累积通路上ACC1、ENO1、PDH-E1α、STP12、SUC1、PSB28、PSBY基因在种子和角果皮中的RNA和蛋白水平的表达量随着种子发育的进程均呈上升趋势,但在不同含油量品种中33DAF或之前的基因表达水平与菜籽含油量呈不完全一致的关系,而43DAF时基因表达水平与菜籽含油量均呈一致的关系,由此推断种子发育后期的基因表达水平与菜籽含油量密切相关,是决定油菜含油量的关键时期。
