HN蛋白糖基化位点缺失的新城疫病毒的生物学特性研究
2020-10-22孙军峰陈琳娜韩宗玺刘胜旺
艾 惠,孙军峰,陈琳娜,李 乐,韩宗玺,刘胜旺
(中国农业科学院哈尔滨兽医研究所 兽医生物技术国家重点实验室,黑龙江 哈尔滨 150069)
新城疫(Newcastle disease,ND)是由新城疫病毒(NDV)感染禽类引起的一种急性、高度接触性传染病,呈世界性分布,严重危害家禽养殖业,能够造成严重的经济损失[1]。NDV遗传进化的变化是一个逐步积累的过程,点突变是NDV变异的主要方式。病毒的不断演化和变异是使NDV的毒力等生物学特性发生变化的一个重要的原因。血凝素-神经氨酸酶(Hemagglutinin-neuraminidase,HN)蛋白是一种II型囊膜蛋白,是NDV的重要免疫原性蛋白和毒力影响因素。研究表明,HN蛋白中和表位(346~353)中的347位氨基酸残基存在E-K的变异,并且含有这种变异的毒株抗原性与传统毒株存在抗原性差异[2]。
N-链接的糖基化修饰作为一种最常见的蛋白质翻译后修饰,病毒囊膜糖蛋白能够利用细胞的这一功能进行修饰,并最终影响病毒蛋白的稳定性、抗原性和生物学功能[3-4]。NDV的HN蛋白含有6个潜在的N-链接的糖基化修饰位点(119、341、433、481、508位和538位残基),分别命名为G1、G2、G3、G4,G5和G6,其中前4个确实发生N-链接的糖基化修饰。据报道,HN蛋白跨膜区突变和N-糖基化缺失影响病毒的吸附、神经氨酸酶活性和融合促进活性[5]。通过反向遗传学技术拯救获得了HN蛋白糖基化位点缺失的突变病毒,进一步研究表明,HN蛋白糖基化缺失能够致使毒力降低[6]。
本研究分离到了1株NDV ck/CH/LHLJ170302,遗传进化分析发现其属于class II类中的基因II型,与LaSota疫苗株亲缘关系较近。基因分子特征分析发现,ck/CH/LHLJ170302株HN蛋白中缺失了433位(G3)的糖基化修饰位点,为探究糖基化位点缺失对ck/CH/LHLJ170302生物学特性的影响,本研究比较分析了ck/CH/LHLJ170302株与LaSota株在生长特性、致病性和抗原性方面的差异,为NDV的遗传变异提供参考数据。
1 材料与方法
1.1 主要试验材料及实验动物 LaSota疫苗株;黑龙江某养鸡场中采集咽拭子样品,分离出1株NDV毒株,命名为ck/CH/LHLJ170302。DF-1细胞由本实验室传代培养;9~11日龄的SPF鸡胚、SPF鸡及鸡红细胞均由中国农业科学院哈尔滨兽医研究所实验动物中心提供。
1.2 主要试剂 体液病毒DNA/RNA小量提取试剂盒,购自Axygen公司;PrimeScriptTMOne Step RT-PCR Kit、Gel Extraction Kit胶回收纯化试剂盒,均购自美国OMEGA公司;pMD18-T载体、ExTaq®DNA Polymerase,均购自宝生物工程(大连)有限公司。
1.3 病毒的纯化 根据文献[7]中的方法步骤,用鸡胚有限稀释法纯化病毒,纯化的病毒-70 ℃保存备用。
1.4 病毒RNA的提取、基因组的扩增及测序 利用体液病毒DNA/RNA小量提取试剂盒提取纯化的ck/CH/LHLJ170302株病毒基因组RNA。利用文献[7]中报道的NDV class II全基因组测序引物序列,用一步法RT-PCR试剂盒扩增ck/CH/LHLJ170302株基因组各片段。回收扩增后的片段并克隆至pMD18-T载体中,转化感受态细胞后挑取单个克隆进行菌液PCR鉴定。同一片段挑取多个阳性克隆,由哈尔滨睿博生物技术有限公司进行测序,取3个以上的相同测序结果作为每个目的片段的序列结果。
1.5 病毒基因分子特征分析 以LaSota疫苗株为参考,根据测序结果拼接获得ck/CH/LHLJ170302株的全基因组序列,比较ck/CH/LHLJ170302株和LaSota株之间各基因的核苷酸、编码的氨基酸的相似性以及F蛋白和HN蛋白的N-链接糖基化位点、半胱氨酸残基及中和表位氨基酸的差异。
1.6 病毒的遗传进化分析 从GenBank中下载19株NDV代表性毒株的F基因编码序列作为参考序列,构建F基因系统进化树。
1.7 病毒的生长动力学特性及致病指数测定 根据文献[7]中的方法,测定ck/CH/LHLJ170302株和LaSota株的鸡胚半数感染量(EID50),以100 EID50的剂量接种9日龄SPF鸡胚,不同时间点收取尿囊液测定病毒滴度,用GraphPad Prism 5软件绘制生长动力学曲线。根据世界动物卫生组织(OIE)致病指数测定标准,测定LaSota株与ck/CH/LHLJ170302 株的鸡胚死亡平均时间(Mean death time,MDT)和1日龄SPF雏鸡脑内接种致病指数(Intracerebral pathogenicity index,ICPI)。
1.8 病毒抗血清的制备 将4周龄的SPF鸡随机分为2组并饲养于负压隔离器中,每组5只;将LaSota株和ck/CH/LHLJ170302株按106EID50的量通过点眼和滴鼻的方式分别进行免疫接种,0.2 mL/只。首次免疫2周后,以同样剂量通过静脉注射途径加强免疫。加强免疫14 d后,扑杀采血,分离血清,将血清置于56 ℃灭活30 min,分装后,于-70 ℃ 保存待用。
1.9 病毒抗原相关性分析 用纯化的病毒和制备的血清在DF-1细胞中进行交叉中和试验以研究ck/CH/LHLJ170302株与LaSota株的抗原性差异,按Archetti方法计算抗原差异系数R[8]。判断标准:R<0.5,表明病毒之间的抗原有较大差异;0.5≤R≤0.67,表明病毒之间的抗原差异较小;0.67≤R≤1.5,表明病毒之间抗原没有差异。
2 结果
2.1 遗传进化及病毒基因序列分析 测序拼接结果显示,ck/CH/LHLJ170302株基因组全长为15 186nt。基因组结构特征与class II类NDV毒株一致。F基因遗传进化分析结果表明,ck/CH/LHLJ170302株属于class II类基因II型病毒,且ck/CH/LHLJ170302 与class II中基因II型LaSota疫苗株亲缘关系最近(图1),进一步比较分析了ck/CH/LHLJ170302株与基因II型LaSota疫苗株各基因编码区的核苷酸序列及推导的氨基酸序列,结果显示,ck/CH/LHLJ170302株与LaSota株的各基因之间均具有较高的核苷酸和氨基酸一致性(表1)。
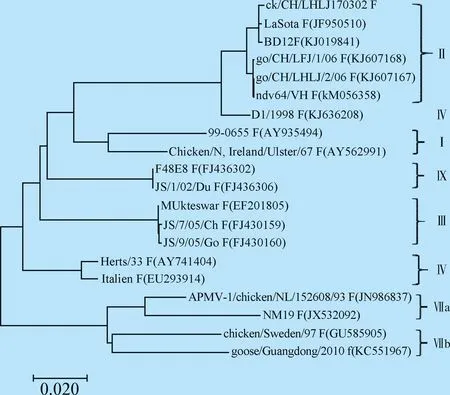
图1 Class II 类 NDV 的 F 基因遗传进化分析Fig.1 The phylogenetic tree of class II NDV strains based on the F gene
2.2F基因序列分析 ck/CH/LHLJ170302株F基因开放阅读框(ORF)长度为1 662 bp,编码553个氨基酸,包含13个半胱氨酸和6个潜在的糖基化位点,均与LaSota株一致。ck/CH/LHLJ170302株F蛋白裂解位点处氨基酸为112G-R-Q-G-R-L117,具有NDV弱毒株的典型特征(表2)。F蛋白中和抗原表位分析发现,ck/CH/LHLJ170302株中和抗原表位的氨基酸与LaSota株中和抗原表位的氨基酸一致。另外,与LaSota株相比,二者F蛋白氨基酸的同源性为99.6%(表1)。上述结果表明,ck/CH/LHLJ170302株与LaSota株的F基因分子特征无明显差异。
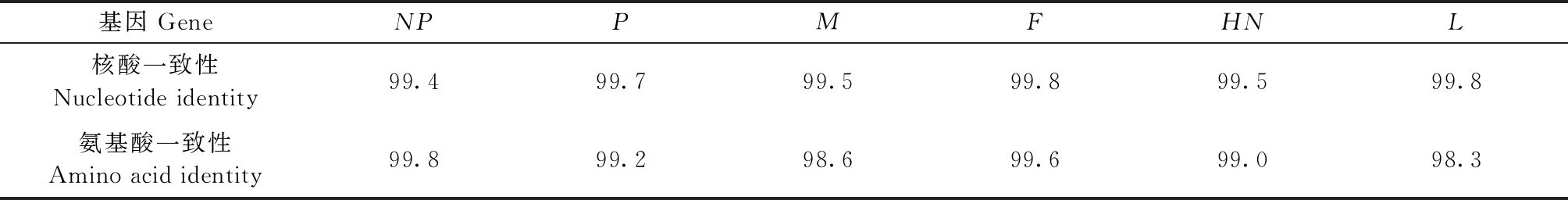
表1 ck/CH/LHLJ170302与LaSota各基因编码区相似性比较Table 1 Comparison of similarity between coding regions of each gene of ck/CH/LHLJ170302 and LaSota (%)
表2 ck/CH/LHLJ170302病毒株与LaSota疫苗株的生物学特性比较Table 2 Comparison of biological characteristics of ck/CH/LHLJ170302 virus strain and LaSota vaccine strain
2.3HN基因序列分析 ck/CH/LHLJ170302株HN基因ORF全长为1 734 bp,编码577个氨基酸,含有12个半胱氨酸残基,与LaSota株一致。但ck/CH/LHLJ170302株的HN基因仅含有5个潜在的糖基化修饰位点,与LaSota株相比,缺失了433位(G3)的糖基化修饰位点。另外,ck/CH/LHLJ170302株HN蛋白中抗原位点的氨基酸序列与LaSota的氨基酸序列没有差异。
2.4 病毒的滴度及致病指数测定 ck/CH/LHLJ170302株和LaSota株的病毒滴度测定结果表明,二者的血凝(HA)滴度和EID50之间没有显著差异。毒力指标测定结果(MDT和ICPI)表明,ck/CH/LHLJ170302株符合NDV弱毒株的标准,与LaSota相比,ck/CH/LHLJ170302和ICPI值有所降低,但ck/CH/LHLJ170302的MDT则提前了近20 h(表2)。
2.5 病毒的生长动力学 将ck/CH/LHLJ170302株和LaSota株接种SPF鸡胚,测定病毒的生长动力学曲线。结果表明,2株病毒均在接种鸡胚96 h后达到最高生长滴度,但ck/CH/LHLJ170302株在24 h、48 h和72 h病毒滴度均显著高于LaSota株(P<0.05)(图2)。
图2 ck/CH/LHLJ170302病毒株与LaSota疫苗株在SPF鸡胚中的生长动力学曲线Fig.2 Growth kinetics curve of ck/CH/LHLJ170302 strain and LaSota vaccine strain in SPF chicken embryo注: 与LaSota组比较,*:P<0.05Note: Compare with LaSota,*:P<0.05
2.6 抗原相关性分析 制备ck/CH/LHLJ170302株和LaSota株的血清后,在DF-1细胞中进行交叉中和试验以评价二者的抗原相关性。结果显示,ck/CH/LHLJ170302与LaSota的抗原差异系数值R为1,表明ck/CH/LHLJ170302与LaSota没有明显的抗原差异。
3 讨论
NDV在家禽中持续存在和流行以及由于点突变的积累而产生的变异病毒持续威胁着家禽养殖业,近年来,由于HN抗原位点变异而产生的NDV抗原变异毒株逐渐增多。此外,NDV HN蛋白中N-链接的糖基化位点对于维持HN蛋白的结构、功能和抗原性具有重要作用[9]。
在先前的研究中,相关研究者利用反向遗传系统获得了缺失HN蛋白4个糖基化位点(G1、G2、G3和G4)以及同时缺失G1和G2两个糖基化位点的突变病毒,进一步研究发现,所有糖基化位点缺失的突变病毒的IVPI值与ICPI值均低于亲本病毒,说明糖基化位点缺失致使毒力降低,尤其G4的缺失和G1、G2的联合缺失能够显著降低NDV的致病性。并且,上述突变病毒与亲本病毒的生长曲线结果表明,突变病毒(rG1、rG2和rG3)在感染后32 h和40 h的滴度明显高于亲本病毒[6]。本研究对ck/CH/LHLJ170302株进行了F基因遗传进化分析与各基因分子特征分析,结果表明,ck/CH/LHLJ170302株属于class Ⅱ类基因Ⅱ型,与LaSota疫苗株的亲缘关系较近,并且发现ck/CH/LHLJ170302株的HN蛋白中天然缺失了433位的糖基化修饰位点(G3)。通过病毒的MDT和ICPI这两项致病指数衡量了ck/CH/LHLJ170302株的毒力,结果表明,ck/CH/LHLJ170302株的MDT为79.2 h,相较于LaSota株的MDT,提前了大约20 h。ck/CH/LHLJ170302株的ICPI为0.175,较LaSota株的ICPI值略微有所降低,上述结果可以表明,ck/CH/LHLJ170302为缓发型弱毒株。另外,生长动力学研究发现,ck/CH/LHLJ170302株的生长速度要高于LaSota株。ck/CH/LHLJ170302株与LaSota株在毒力和生长特性上的差异是否是由于ck/CH/LHLJ170302株HN蛋白中糖基化修饰位点的缺失造成的需要进一步探究。此外,鉴于ck/CH/LHLJ170302株的生长速度高于LaSota株,推测ck/CH/LHLJ170302株可能是LaSota株在鸡群中长期存在并适应形成的突变毒株,这需要进一步的证实。
本研究发现了HN蛋白糖基化位点天然缺失的NDV,并且其生物学特性发生了一定程度的变化,表明需要加强对NDV的流行病学监测和研究,为我国NDV的遗传变异防控提供参考数据。
