稻虾共作模式对稻田二化螟的影响
2020-10-20肖求清汪金平曹凑贵
郭 灿 肖求清 汪金平 曹凑贵
(华中农业大学 湖北武汉 430070)
水稻是世界三大主要粮食作物之一,也是中国最重要的粮食作物[1],我国65%以上人口以稻米为主食[2],但长期单一种植水稻使稻田生物多样性单一化,生态稳定性变差,水稻病虫害加重,并且长期过量使用农药、化肥严重污染环境、使土壤板结等弊端[3]。稻虾共作模式是应用食物链理论、生态位理论和种间互利共生理论,将水稻种植和小龙虾养殖有机结合的新型生态农业模式[4]。本试验以区域调查和大田控制试验结合,探讨稻虾共作模式对稻田二化螟产生的影响。
1 材料与方法
1.1 试验设计
2015 年和2016 年分别在潜江市后湖农场、白鹭湖农场、积玉口镇选择具有代表性的中稻单作模式(CK)和稻虾共作模式(RC)稻田各3 块,调查二化螟冬前、冬后基数(幼虫、蛹和成虫)、第二代二化螟幼虫数、成虫数和第三代二化螟幼虫数。2016 年调查二化螟冬后基数的所有样地与2015 年冬前基数样地一致。所有样地都选择移栽田。
2016 年在潜江市后湖农场选择中稻单作模式(CK)和稻虾共作模式种养年限1年、2年、5年和10 年(RC1、RC2、RC5 和RC10)各3 块稻田作为样田,共15 块,用于调查第三代二化螟幼虫数。这些田块位于同一村,同一天机械插秧,栽插密度、水稻品种统一,水肥农药等管理基本一致。
1.2 测定方法
2015年水稻收割后,调查稻田二化螟冬前基数,2016 年4 月份,调查冬后基数。按棋盘式取样方法[5],每块稻田选10 个点,每点至少取10 蔸水稻残株,现场剥查稻桩内二化螟越冬幼虫的数量[6],每块稻田至少剥查100 蔸稻茬。记录剥查稻茬蔸数、每蔸稻茬中二化螟越冬幼虫数(包括蛹数和蛹壳数),测量每块稻田栽插密度,计算单位面积(hm2,下同)二化螟虫量和越冬死亡率。计算公式如下:

第二代二化螟幼虫调查:2016 年7 月份,在潜江市后湖农场、白鹭湖农场、积玉口镇选择具有代表性的CK 稻田和RC 稻田各3 块调查第二代二化螟幼虫数,记录调查稻田栽插密度、水稻蔸数、总穗数、枯心穗数和二化螟幼虫数(包括蛹数和蛹壳数),算出每hm2二化螟幼虫数和螟害枯心率,计算公式如下:

第二代二化螟成虫调查:2016 年从7 月1 日开始按每hm2田放30个二化螟性诱捕器,诱捕器底部距离水稻顶叶部10 cm~15 cm[7]。诱捕器由北京中捷四方生物科技公司生产,由白色钙塑板、粘虫胶片和诱芯三部分组成。每2d记录一次数据,记录完数据后换粘虫胶片,每15d换一次诱芯,诱芯为中科院动物所二化螟诱芯B[7-8]。每2d更换二化螟性诱捕器摆放地点,直到诱捕的二化螟成虫开始连续下降。7 月3 日记录第一次数据,7 月29 日记录最后一次数据,共记录数据14次。
第三代二化螟幼虫调查:2016年9月底~10月初,调查稻田第三代二化螟幼虫,每块稻田至少剥查100 蔸水稻,记录每蔸水稻二化螟幼虫数(包括蛹数和蛹壳数),计算出每hm2虫量。
1.3 数据统计分析
使用Microsoft Excel 2016 进行数据处理,Statistix 8.0 软件进行统计分析。
2 结果
2.1 二化螟越冬基数和死亡率
表1为不同调查地点不同类型稻田二化螟越冬基数及越冬死亡率。由表1可知:后湖农场、白鹭湖农场、积玉口镇三个地点RC 稻田二化螟冬前基数分别比CK 稻田少53.1%、59.6%、45.4%。白鹭湖农场与其他两个地点差异比较大,说明二化螟冬前基数存在区域性,与陈丽媛[5]等2013 年研究结果一致。三个地点CK稻田二化螟冬前基数和冬后基数都极显著的高于RC稻田二化螟冬前基数,说明RC 能有效控制第二年稻田内二化螟虫源;三个地点CK 稻田二化螟幼虫越冬死亡率分别为22.7%、23.8%、15.1%,说明二化螟越冬幼虫抗寒性比较强,大部分幼虫经过一个冬天还存活;三个地区的RC稻田二化螟冬后基数全为0头/hm2,可能是RC 模式需要经过冬、春季覆水或越冬小龙虾取食灭绝了稻田内二化螟,从而影响后期二化螟世代的发育进度,RC 稻田二化螟虫源来自周边稻草或为其他稻田迁移过来。
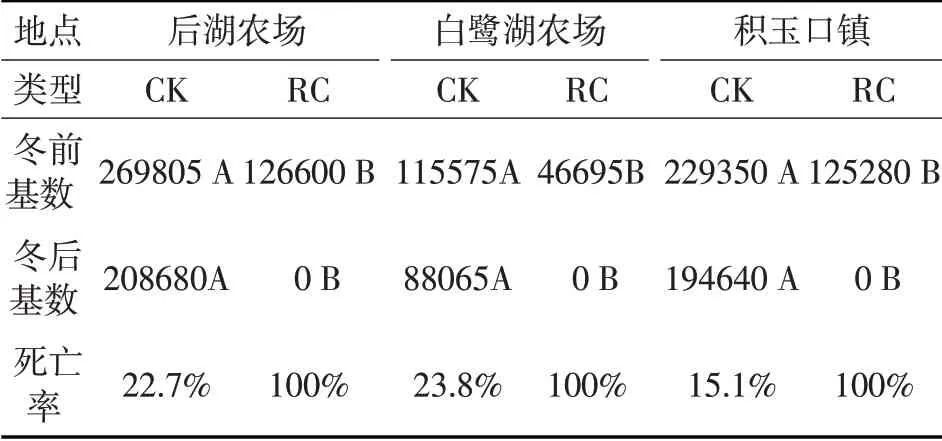
表1 不同地点稻虾共作模式二化螟幼虫越冬基数(头/hm2)
2.2 第二代二化螟幼虫数量
图2为不同调查地点中稻单作和稻虾共作模式的第二代二化螟幼虫数量比较,分析图2可知:后湖农场、白鹭湖农场、积玉口镇CK稻田第二代二化螟幼虫数量都极显著的高于RC 稻田,说明相比CK,RC 极显著减少稻田第二代二化螟幼虫数量。

图2 不同地点的稻虾共作模式第二代二化螟幼虫数
2.3 螟害枯心率
图3 为不同调查地点,不同类型稻田枯心率比较。分析图3可知:CK稻田平均枯心率分别为1.50%、1.19%、1.17%,RC稻田平均枯心率分别为1.07%、0.78%、0.83%。后湖农场、白鹭湖农场CK稻田枯心率显著高于RC稻田,积玉口镇CK稻田枯心率高于RC稻田,但不显著。CK稻田平均枯心率为1.29%,RC 稻田平均枯心率为0.89%,比CK 稻田枯心率减少31.0%,说明RC 能一定程度上控制稻田螟害枯心率。

图3 不同地点稻虾共作和水稻单作稻田螟害枯心率
2.4 第三代二化螟幼虫数
图4为2016年后湖农场不同RC年限稻田的第三代二化螟幼虫数量。分析图4可知:CK稻田第三代二化螟幼虫数量为553875头/hm2,极显著高于其他所有RC年限稻田;所有RC稻田第三代二化螟幼虫平均数量为156090 头/hm2,低于CK稻田71.82%,说明相比CK 稻田,RC 稻田极显著降低第三代二化螟数量。RC1、RC2、RC5、RC10稻田第三代二化螟幼虫数量分别为166440 头/hm2、154440 头/hm2、156480 头/hm2、146985 头/hm2,数值之间差异不显著,说明稻虾共作模式不同种养年限对稻田第三代二化螟幼虫数量没有明显影响,只要是RC模式,不管RC1还是RC10,控制二化螟为害的效果一样。
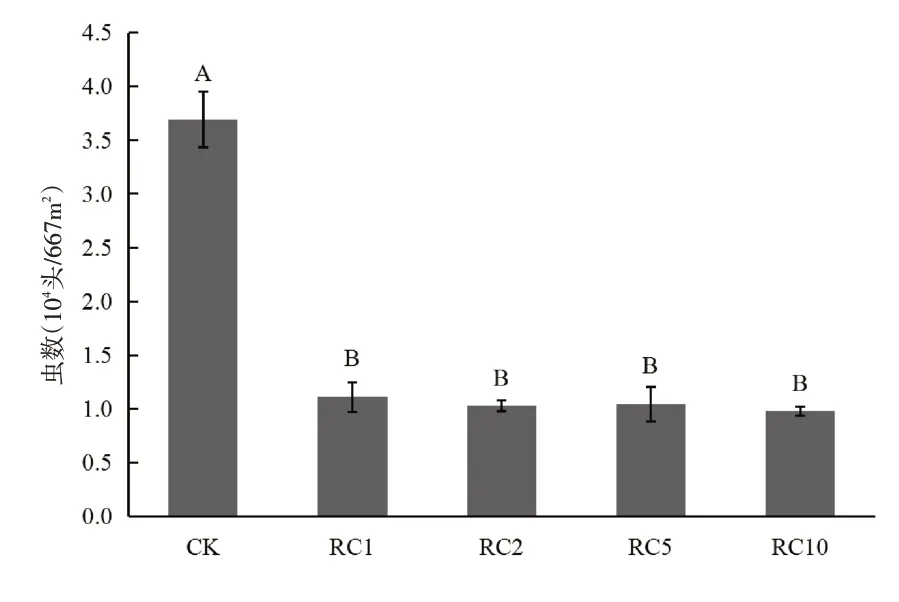
图4 不同稻田年限的第三代二化螟幼虫数
2.5 二化螟成虫数
图5 显示2016 年后 湖农场RC 稻田 和CK 稻田第二代二化螟成虫(蛾)高峰期。分析图5 可知:RC 稻田蛾的高峰期比CK 稻田晚10 d。因此,建议RC稻田二化螟药物防治时期相比CK稻田可以推迟10 d 左右;CK 稻田蛾高峰值为199.95 只/hm2,RC 稻田 蛾高 峰值为130.05 只/hm2,RC 稻田低于CK 稻田34.96%,说明对比CK稻田,RC稻田能显著降低第二代二化螟蛾数,从而减少二化螟卵量,控制第三代二化螟数量。

图5 二化螟性诱捕器诱捕二化螟蛾数
3 结论与讨论
二化螟幼虫主要在稻秆基部活动,其在常规稻的垂直分布中,40%以上越冬幼虫停留在离地10cm 以下的稻秆中,约80%停留在20cm 以下的稻秆中[10]。二化螟幼虫在食料不足或水稻生长受阻时有转株为害的习性[11]。稻虾共作模式中水稻为中稻,一般六月份或以后移栽,而第一代二化螟幼虫为害时期发生在五月份,为害早稻,因此稻虾共作模式稻田不需要调查第一代二化螟幼虫为害。田间调查发现二化螟在同一块稻田存在明显的田间聚集分布。
在本研究中,相比中稻单作模式,稻虾共作模式推迟二化螟发育进度,极显著降低二化螟为害程度,有效抑制二化螟带来的虫害。主要原因可能是稻虾共作模式下冬、春季覆水,田面长期保持50cm 以上水层,使越冬二化螟幼虫缺氧窒息死亡,减少第二年稻田二化螟虫源,改变后期二化螟世代发生为害,从而影响二化螟周年发生规律;另外一个原因可能是小龙虾在冬季覆水时取食二化螟幼虫,影响二化螟成虫产卵量,也能一定程度减轻二化螟为害。
