重组伪狂犬病病毒的研究进展
2020-10-17张传健刘娅梅侯继波王继春
张传健,刘娅梅,侯继波,王继春
(1.江苏省农业科学院 动物免疫工程研究所,南京 210014;2.江苏省农业科学院 国家兽用生物制品工程技术研究中心,南京 210014;3.江苏省动物重要疫病与人兽共患病防控协同创新中心,扬州 225009)
伪狂犬病病毒(Pseudorabies virus,PRV)属于疱疹病毒科甲型疱疹病毒亚科水痘病毒属,能感染猪、牛、羊和犬等多种家畜,引发伪狂犬病。猪伪狂犬病对我国养猪业危害极大,可造成母猪发热、精神不振、进而发生流产或产木乃伊胎、死胎和弱仔。仔猪高热、厌食、呼吸困难、神经症状、并伴随着腹泻,症状持续5~10 d,发病和死亡率可达100%。成年猪通常为隐性感染。研究者通过基因工程技术构建的gE、gE/gI或TK/gE/gI基因缺失株能够有效地抵抗强毒的攻击,是构建猪用重组疫苗的理想载体[1-2]。利用PRV基因缺失株为载体构建表达异源抗原基因重组病毒,对宿主安全可靠,既能对伪狂犬病提供保护,又能产生针对异源保护性抗原的特异性免疫反应,可达到一针多防的效果。本文从PRV基因组结构与特征、重组PRV构建方法、PRV缺失株的构建与应用以及PRV活载体疫苗的研究进展四个方面进行综述。
1 PRV基因组结构与特征
PRV为双链DNA病毒,属于甲型疱疹病毒亚科水痘病毒属,其基因组约为150 kb,GC含量高达74%,可分为独特长区段(unique long,UL)、内部重复区段(internal repeat sequence, IRS)、独特短区段(unique short,US)以及末端重复区(terminal repeat sequence,TRS)。随着对PRV的基因组的解析,各基因的定位已基本明确。UL区被鉴定的基因包括gB、gC、gK、gH、主要衣壳蛋白基因、碱性核酸酶基因、核苷酸还原酶基因、胸腺激酶基因(thymidine kinase,TK)和DNA多聚酶基因等;IRS区的基因主要有立即早期基因和RSP40基因等;US区的基因包括gD、gE、gG、gI以及蛋白激酶基因等。PRV基因可编码结构蛋白、免疫调节蛋白、毒力相关蛋白、病毒的转录因子、病毒复制及释放相关蛋白等70多种蛋白。其中包括16种囊膜蛋白,分别是11种糖蛋白(gB、gC、gD、gE、gG、gH、gI、gK、gL、gM和gN)、4种跨膜蛋白(UL20、UL43、US9和可能包括UL24)和II型膜蛋白。囊膜蛋白主要涉及病毒进出细胞以及在细胞间的传播,还调节免疫反应,促进合胞体形成。糖蛋白作为病毒粒子和感染细胞的表面成分,对宿主免疫防御起重要作用。其中,糖蛋白gB、gC和gD是PRV主要的保护性抗原,刺激机体产生中和抗体和细胞免疫反应。研究发现PRV复制非必需基因超过一半,这些基因通常是毒力相关基因,毒力相关基因的缺失可降低PRV的毒力而不影响病毒的生长,因此,PRV是异源基因插入的理想载体。
2 重组PRV构建方法
随着生物技术的发展,针对疱疹病毒进行DNA修饰(点突变、基因敲除和基因插入)的技术手段已十分成熟。构建重组PRV的关键是PRV基因修饰后能够在宿主细胞内包装出具有感染性的重组PRV粒子。目前用于构建PRV重组病毒技术方法包括传统同源重组技术、细菌人工染色体(bacterial artificial chromosome,BAC)技术和CRISPR/Cas9介导的基因编辑技术。传统同源重组方法重组效率较低,重组病毒纯化过程费时费力。BAC技术是研究疱疹病毒有力的工具。PRV感染性克隆可在大肠杆菌中以单拷贝或极低拷贝质粒的形式进行复制,不易发生变异,还能通过Red重组对BAC DNA进行遗传操作,效率高,成本低,操作精度高,将BAC DNA转染宿主细胞可成功拯救出重组病毒。利用BAC技术构建的重组病毒不需要在宿主细胞传代和筛选,简化了重组病毒的构建。CRISPR/Cas9介导的基因组定点编辑技术是近些年研究的热点,结合同源重组和非同源重组对PRV进行基因的插入、缺失和替换。CRISPR/Cas9技术的应用使重组PRV构建的速度更快、效率更高,此外可以对PRV基因组多个位点同时进行修饰,应用前景广阔。
3 PRV基因缺失株的构建与应用
猪伪狂犬病对部分养猪大国危害极大,造成严重的经济损失,病猪和带毒猪是最重要的传染源,PRV可以在猪体形成潜伏感染,对猪群造成持续威胁,是控制该病的难点,基因缺失疫苗配合鉴别诊断技术推动了该病在很多国家家养猪群中的净化。PRV基因缺失疫苗优点在于毒力减弱,返强可能性极低,保持较好的免疫原性。大多数缺失株不能侵入神经系统,难以建立潜伏感染。第一代基因工程PRV缺失株为TK单基因缺失株。1984年,Kit等[3]以PRV BUK为亲本株构建了TK单基因缺失株PRV BUK-dl3株,试验表明,PRV BUK-dl3株对猪只安全,攻毒保护率较高。该毒株是世界上首个获得批准使用的基因工程疫苗,1986年在美国上市。我国研究者陈焕春等[4]和王琴等[5]分别构建了PRV Ea和Ra TK基因缺失株,研究表明两株缺失株体外生长特性稳定。第二代基因工程PRV缺失株不仅缺失TK基因,还缺失复制非必需的糖蛋白基因(TK/gE缺失株、TK/gG缺失株、TK/gC缺失株)或者添加报告基因,主要用于疫苗毒株和野生毒株的鉴别诊断[6-8]。Bartha-K61株是一种gI/gE双基因缺失弱毒疫苗,缺失部分包括整个gE基因以及部分gI基因,毒力基因的缺失大大提高该疫苗的安全性,并且能够有效地保护免疫动物。Bartha-K61株的推广应用对很多国家家养猪群PRV的净化发挥了重大作用[9]。1970年,Bartha-K61株被引入我国并广泛使用,使我国猪伪狂犬病得到较好的控制。2000年之后,许多猪场建立了PRV gE抗体阴性猪群,不少猪场基本实现PRV的净化。然而,2011年以来,一种PRV变异株在我国暴发流行,变异株不仅造成仔猪发病死亡,母猪流产,同时对生长猪和育肥猪致病力较强,甚至引起死亡[10-11]。Bartha-K61株不能提供完全的保护力(阻止排毒的效力严重不足)[12-14]。目前,国内多个团队研究发现变异株缺失株(gE/gI、gE/TK或者TK/gE/gI)能够有效地抵抗变异株攻击,如表1所示。本课题组在2012年分离鉴定了PRV变异株AH02LA株,研究发现该变异株引起4~5周龄仔猪和9~10周龄生长猪100%发病或死亡,感染后2~3 d,4~5周龄仔猪出现神经症状、呼吸症状、腹泻,9~10周龄生长猪出现明显的呼吸症状,表明该变异株毒力增强[15]。之后本课题组通过细菌人工染色体技术构建了AH02LA株gE和TK双基因缺失株,研究发现该缺失株接种PRV阴性断奶仔猪后对变异株AH02LA株攻击产生完全的保护[13]。

表1 不同PRV变异株缺失株对仔猪免疫效力的研究Table 1 Study on protection efficacy induced by gene deleted PRV variants in piglets
gB、gC和gD是PRV的主要保护性抗原,Yu等[22]以Bartha-K61株为载体,利用CRISPR/Cas9技术将Bartha-K61株的gB基因替换为变异株JS-2012株的gB基因,制备的灭活疫苗接种小鼠可对变异株提供80%的保护。本研究团队以Bartha-K61株为载体,利用BAC系统将Bartha-K61株的gC基因和gD替换为变异株AH02LA株的gC基因和gD基因,制备活疫苗接种断奶仔猪可对变异株AH02LA株攻击产生完全的临床保护,降低排毒率或减少排毒量、缩短排毒时间,非常适用于我国猪场中PRV变异株的净化[14]。PRV变异株缺失株的构建与应用,为PRV变异株活载体疫苗的构建奠定了基础。
4 PRV活载体疫苗的研究进展
随着生物技术的发展,基因工程病毒活载体疫苗成为新一代兽用疫苗研究的热点。疱疹病毒基因组大,含多个复制非必需基因和非编码区,对外源基因的承载力强,适合用于载体疫苗的构建。研究发现以疱疹病毒为载体构建的重组病毒能有效地表达外源基因,快速产生高效的体液免疫和细胞免疫,应用效果显著。此外,活载体病毒的生产工艺简单,成本低廉,使用方便。
PRV属于疱疹病毒,其基因缺失活疫苗被广泛用于猪伪狂犬病的预防,因此以PRV缺失株为载体,插入猪病病原的保护性抗原构建活载体疫苗有预防多病的潜力。国内外在这方面进行了诸多探索。
4.1 猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory syndrome,PRRSV)/PRV重组病毒Qiu等[23]将PRRSV GP5基因插入到Bartha-K61株TK基因的等位位点,构建了表达GP5基因的重组病毒rPRV-GP5,免疫仔猪没有检测到PRRSV抗体,PRRSV CH-1a强毒株攻击后,免疫仔猪出现温和的体温反应,轻微的肺脏和肾脏病理变化以及短期的病毒血症。为了提高rPRV-GP5的保护效率,江云波[24]等将改造过的PRRSV GP5m+基因插入PRV Ea基因缺失疫苗TK-/gE-/LacZ+基因组中,构建重组病毒rPRV-GP5m,免疫小鼠产生PRRSV中和抗体和细胞免疫反应水平优于rPRV-GP5。之后该团队将GP5m基因和膜蛋白M基因插入PRV TK-/gE-/LacZ+基因组中,构建重组病毒rPRV-GP5m-M,免疫小鼠诱发抗PRV特异性抗体,并能抵抗PRV致死性的攻击,同时产生高水平PRRSV特定的中和抗体和淋巴细胞增殖反应。免疫仔猪后,与PRRSV灭活苗相比,rPRV-GP5m-M诱导更高水平的PRRSV特定中和抗体和淋巴细胞增殖反应。PRRSV强毒攻击后,rPRV-GP5m-M接种仔猪组织内病毒载量降低,病毒血症时间缩短[25],该重组病毒的构建为PRRSV/PRV活载体疫苗的研究奠定了基础。
4.2 猪圆环病毒II型(Porcine circovirus type 2,PCV2)/PRV重组病毒Ju[26]等将PCV2融合蛋白ORF1和ORF2基因插入PRV Ea株gE和gI基因的等位位点,构建了共表达ORF1和ORF2基因的重组病毒rPRV-PCV2,免疫小鼠和猪后,诱导PRV和PCV2特定抗体以及PCV2淋巴细胞增殖反应。之后该研究团队构建了表达ORF2基因的重组病毒PRV Tk-/gE-/ORF2+,免疫4周龄仔猪诱发抗PRV和PCV2体液免疫反应,免疫后3周仍能检测到PCV2特定淋巴细胞增殖反应。Zheng等[27]构建了共表达ORF2和IL-18基因的PRV重组病毒PRV-ORF2-IL18,免疫小鼠诱导高水平的抗PRV和PCV2抗体以及PCV2特定淋巴细胞增殖反应(CD3+,CD4+和CD8+T淋巴细胞数量增加),且能有效地抵抗PRV强毒株的攻击,减少PCV2强毒株攻击后组织的病毒载量,以上研究结果表明该重组毒具有预防PCV2和PRV的巨大潜力。
4.3 口蹄疫病毒(Foot-and-mouth disease virus,FMDV)/PRV重组病毒Ding等[28]构建了表达FMDV VP1基因的PRV重组病毒rPRV-VP1。免疫猪抗FMDV抗体水平低于FMDV商品疫苗,减轻FMDV强毒攻击后的临床症状。为了提高重组病毒的免疫效率,将含有FMDV VP1和SV40 polyA基因的表达盒插入PRV弱毒株gG基因的等位基因,构建重组病毒PRV-P1,免疫猪后,重组病毒组产生高水平抗PRV和FMDV抗体,激活抗FMDV细胞毒性T淋巴细胞反应,FMDV强毒株攻击后,仔猪存活率为3/5[29]。之后Zhang等[30]将FMDV的衣壳前体多肽P12A和非结构蛋白3C基因插入到PRV弱毒株gI和gE基因的等位基因,构建了重组病毒PRV-P12A3C,免疫6周龄仔猪诱发了高水平抗FMDV中和抗体和特定淋巴细胞增殖反应,FMDV强毒株攻击后,FMDV灭活苗产生100%的保护,PRV-P12A3C诱导60%的保护。由于目前FMDV/PRV重组病毒效果不佳以及FMDV血清型众多,FMDV/PRV重组疫苗尚需进一步研究。
4.4 猪流感病毒(Swine influenza virus,SIV)/PRV重组病毒Tian等[31]构建了表达H3N2 HA基因的PRV基因重组病毒rPRV-HA,免疫小鼠三周后诱发血凝抑制抗体。H3N2强毒攻击后,免疫小鼠没有检测到H3N2强毒,肺部发生温和病变。Walter Fuchs等将H1N1密码子优化的HA或NA基因插入到PRV Bartha-K61株gG基因的等位基因,构建重组病毒PrV-BaMI-synH1和PrV-BaMI-synN1。PrV-BaMI-synH1和PrV-BaMI-synN1分别或者混合免疫7周龄生长猪后,诱导B细胞增殖反应和H1N1特定抗体,H1N1强毒攻击后,PrV-BaMI-synH1或PrV-BaMI-synN1免疫猪无临床症状,排毒量显著减少。此外,研究结果表明PrV-BaMI-synH1保护效率优于PrV-BaMI-synN1。滴鼻接种PrV-BaMI-synH1两次能够阻止H1N1强毒株的接触性感染[32-33]。
4.5 猪瘟病毒(Classical swine fever virus,CSFV)/PRV重组病毒Ben等[34]构建了表达E2基因的PRV gD和gE缺失的重组病毒,免疫猪有效地抵抗PRV强毒株和CSFV的攻击。Wang等[35]将CSFV E2基因插入PRV TJ株gE和gI基因的等位位点,构建重组病毒rPRVTJ-delgE/gI-E2,PRV变异株和CSFV强毒株攻击猪后,rPRVTJ-delgE/gI-E2免疫组快速诱导高水平抗PRV和SCFV中和抗体,产生100%的保护[35]。Lei等[36]将CSFV E2 基因插入PRV TJ基因缺失病毒PRV TJ-delgE/gI/TK内,构建重组病毒rPRVTJ-delgE/gI/TK-E2,免疫猪没有发生临床症状和排毒,诱导高水平抗PRV和SCFV中和抗体,且能有效地抵抗PRV TJ株和CSFA Shimen株的攻击,为CSFV/PRV双价疫苗的研究奠定了基础。
4.6 日本乙型脑炎病毒(Japanese encephalitis virus,JEV)/PRV重组病毒Xu等[37]构建了表达JEV NS1基因的PRV重组病毒rPRV-NS1。105PFU重组毒接种小鼠和仔猪后诱导JEV特定的细胞免疫和体液免疫反应,有效地抵抗PRV强毒Ea株的攻击。Qian等[38]将JEV I型prM和E基因插入PRV弱毒疫苗株PRV TK-/gE-/lacZ+内,构建重组病毒PRV TK-/gE-/PrM-E+。免疫小鼠产生抗PRV和JEV ELISA抗体。JEV SX09S-01强毒株攻击后,免疫组死亡时间延长,死亡率降低。
4.7 猪细小病毒(Porcine parvovirus,PPV)/PRV重组病毒Chen等[39]将PPV VP2基因插入到PRV SA215株 gI和gE基因的等位基因,构建重组病毒PRV SA215/VP2,免疫仔猪后,重组病毒组诱导抗PRV和PPV体液免疫反应,完全抵抗PRV强毒的攻击。免疫母猪后,诱导PPV特定抗体并且显著减少PPV强毒株攻击的流产率。
普遍认为活载体疫苗比其他新型疫苗更有发展潜力,但获得能够商品化的重组活载体疫苗并非易事,目前PRV活载体疫苗没有一个进入生产应用中。启动子和插入位点的选择是制约PRV活载体疫苗发展的重要因素。合适的插入位点不仅可以使异源基因稳定表达,还不影响病毒的生长特性。如表2所示,目前报道的重组PRV的插入位点主要包括gG等位位点、PK和gG之间、gI等位位点、gI和gE的等位位点和TK的等位位点。研究发现外源基因插入以上几个位点能够成功稳定的表达,然而一些研究并未提供重组病毒的体外生长动力学的数据,外源基因插入以上几个位点是否影响病毒的复制尚需鉴定。合适的插入位点选择将是今后的研究重点。报道中表达外源基因的启动子主要选用gG、HCMV、MCMV和CAG(包括HCMV增强子和鸡β-actin启动子)。Alexandra等[40]使用3种不同的启动子(HCMV、MCMV和CAG),构建了3种表达ASFV pE199L基因重组PRV病毒,发现CAG作为启动子pE199L表达水平最高。相对于单一的启动子,复合启动子的使用可能会增加外源基因的表达量。
5 PRV重组病毒的应用前景
针对我国出现的PRV变异株,国内多个团队通过基因工程技术构建多株PRV变异株基因缺失活疫苗,尽管活疫苗在安全性上可能存在少许不足,但合理地使用这些疫苗可产生良好的效果,为我国PRV的净化奠定坚实的基础。此外,国内外很多团队构建多种表达异源病毒抗原基因的PRV重组病毒免疫动物可诱导产生体液免疫和细胞免疫,但报道的免疫效力有高有低,差别很大,目前相关疫苗的应用较少。分析效果的影响因素主要包括外源保护性基因的选择、目的抗原基因插入位点和表达、载体免疫与目的抗原免疫的竞争等。PRV基因组庞大,有很多可供选择的复制非必须区,如果选择合适的位点插入猪病的保护性抗原基因可以达到接种一种疫苗预防多种疾病的效果,既节省成本提高效率,又减少免疫次数,减少应激,提高经济效益,插入位点选择的基本要求是外源基因插入后不影响载体病毒增殖能力和免疫原性,且外源基因插入后能保持稳定,不发生丢失,大多报道的研究选择非必需基因进行替换插入外源基因表达盒,虽然替换的是非必需基因,但可能对病毒的复制和稳定性产生影响,因此,选择基因开放阅读框下游的非编码区作为插入位点,以避免对上下游基因的影响,应是PRV重组活载体的研究内容之一。此外,启动子选择的好坏制约外源基因的表达水平,选择合适的启动子可增加目的抗原基因的表达水平,进而提高重组病毒的免疫效力,报道中一般选择MCMV IE、SV40和Beta-actin等强启动子来增强外源基因的表达,在接下来的研究中可以将这些启动子进行复合或者引入甲病毒复制子以进一步提高外源抗原的表达量。由于猪群广泛接种PRV疫苗,PRV母源抗体的干扰也是PRV载体活疫苗在临床上应用的重大障碍,因此,如何克服PRV母源抗体的干扰也是PRV载体活疫苗在生产中能得到广泛应用的关键之一,通过矿化方法或壳聚糖等对载体病毒进行包裹有望一定程度克服母源抗体干扰,还有通过滴鼻、喷鼻或雾化等途径进行呼吸道接种也能一定程度避免母源抗体的干扰作用,是提高PRV载体活疫苗临床应用价值的方法,值得进一步研究。另外,由于PRV的宿主谱非常广,在其他宿主上没有母源抗体干扰的问题,通过构建PRV必需基因缺失株载体与配套表达细胞系生产复制缺陷型的载体疫苗株也是对除猪以外的家畜开发以PRV为载体的新型疫苗的思路之一。综上所述,为开发出能在生产中应用的PRV活载体疫苗,以上因素都是我们今后研究的重点。
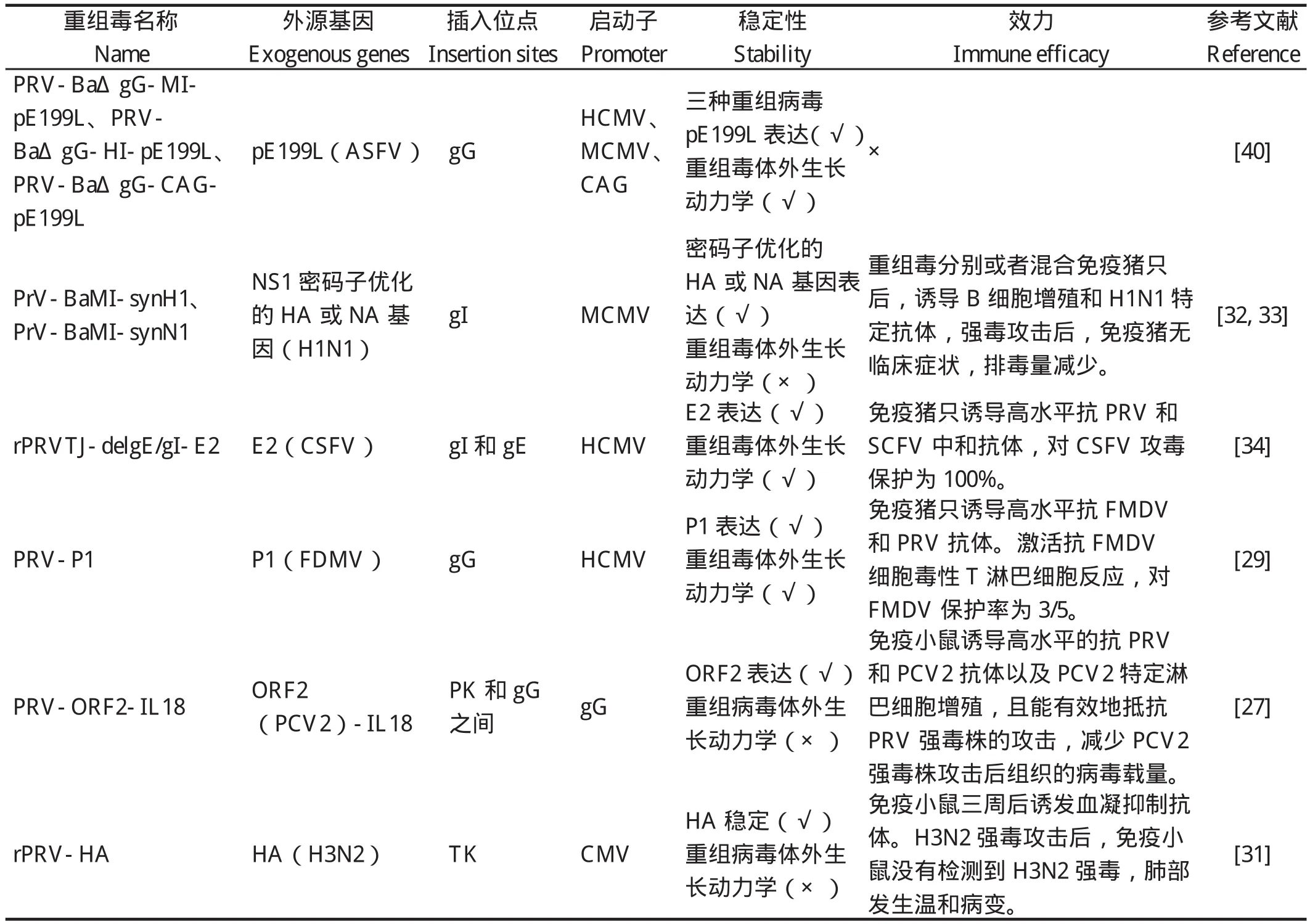
表2 重组PRV的基本信息Table 2 General information of recombinant PRVs
