冷藏鱼片中优势腐败菌的分离鉴定
2020-09-23赵欣宇孙卫青熊光权吴文锦丁安子
赵欣宇,孙卫青,熊光权,乔 宇,吴文锦,李 新,丁安子,汪 兰,*
(1.湖北省农业科学院农产品加工与核农技术研究所,湖北省农业科技创新中心农产品加工研究分中心,湖北武汉 430064;2.长江大学生命科学学院,湖北荆州 434025)
随着生鲜电商、连锁餐饮、中央厨房等新零售业态的崛起,水产品的消费从鲜活逐渐转向预制调理加工。预制水产品在冷藏过程中会腐败变质,主要是由于水产品水分和蛋白质含量高,微生物容易大量繁殖。现有研究主要集中在优势腐败菌的分离鉴定和致腐能力的研究,特别是希瓦氏菌(Shewanella)和假单胞菌(Pseudomonasspp.)[1-3]。而气单胞菌也是鱼、贝、虾蟹类等水产品的优势腐败菌之一。钱韻芳等[4]从气调包装凡纳滨对虾中分离纯化了气单胞菌,并对其致腐能力进行了研究,结果表明气单胞菌能使凡纳滨对虾挥发性盐基氮含量(Totalviablenitrogen,TVB-N)、三甲胺氮(Trimethylaminenitrogen,TMA-N)含量升高,从而产生异臭味物质;李婷婷等将无菌鲢鱼块接种温和气单胞菌,结果表明气单胞菌能使鱼肉TVB-N、TMA含量升高,细菌蛋白酶能使鱼肉蛋白质、脂肪等分解成大量小分子物质,产生大量离子从而使其电导率升高,Ca2+-ATPase活性降低等造成鱼肉的蛋白质等营养物质劣变[5-9]。
斑点叉尾鮰(Leiocassislongirostris)是中国主要的淡水经济鱼类之一,受到广大消费者的青睐。近年来学者对冷藏鲢鱼、冷藏鲟鱼、冷藏金枪鱼等[3,5,9-12]的优势腐败菌及相关研究较多,但对冷藏鮰鱼的优势腐败菌的研究较少。本文以斑点叉尾鮰为原材料,将新鲜鮰鱼于4 ℃无菌冷库中取肉后真空包装,放置于4 ℃冰箱中贮藏7 d。从腐败后期鱼肉中分离出优势腐败菌并对其进行细菌鉴定,以期为冷藏鮰鱼片保鲜技术的研究提供科学依据。
1 材料与方法
1.1 材料与仪器
新鲜鮰鱼 湖北省武汉市华南海鲜批发市场;麦康凯琼脂培养基、血平板培养基、营养琼脂培养基 青岛海博生物技术有限公司;细菌生化微量鉴定 广东环凯微生物科技有限公司;2×Taq PCR Master Mix细菌基因组DNA提取试剂盒、DNA胶回收试剂盒 天一辉远生物科技有限公司;PCR板 爱思进生物技术(杭州)有限公司(AXYGEN)。
YX280型高压蒸汽灭菌锅 上海三中医疗器械有限公司;DL-CJ-2NDH型超净工作台 北京东联哈尔仪器制造有限公司;BPC-150F型生化培养箱 上海一恒科学仪器有限公司;WD-9406型胶片观察灯 北京六一仪器厂;Mikro120 台式离心机 广州市华粤行仪器有限公司;Thermal Cycler2720型PCR仪 基因有限公司Applied Biosystem;DYY-6C型电泳仪 北京六一仪器厂;Gel DocTM XR+型紫外分析仪 美国伯乐BIO-RAD;3730XL型ABI 3730 基因有限公司Applied Biosystem。
1.2 实验方法
1.2.1 优势腐败菌的分离纯化 在4 ℃无菌冷库中将鮰鱼宰杀,去掉其头、皮、刺及内脏,用无菌水将其冲洗干净,手工取侧线上方、背鳍附近的白肉约20 g,真空包装后置于4 ℃冰箱中储藏7 d。参照GB 4789.2-2016[13]的方法将适量鱼肉于超净工作台中剪碎于装有适量的生理盐水的锥形瓶中摇匀,制成菌液。将菌液依次进行10倍梯度稀释,取适当梯度的菌液1 mL于血平板培养基、营养琼脂培养基中,培养腐败菌,温度为37 ℃,时长24 h。观察并挑选各个平板上的优势腐败菌用平板划线法[14]纯化至形态特征稳定的菌种并编号。
1.2.2 革兰氏染色及生化鉴定 将纯化菌种进行革兰氏镜检,观察其形态。无菌条件下将纯化菌落按照生化微量鉴定管说明书进行氧化酶、非麦芽糖、乙酰胺、硝酸盐还原、西蒙氏柠檬酸盐、DNA等生化鉴定[15-16]。
1.2.3 细菌DNA提取 采用细菌基因组提取试剂盒上方法提取细菌总DNA:取适量菌体,加入2 mL离心管中;加入567 μL的TE缓冲液,反复吹打使之重新悬浮。然后加入30 μL10%SDS和15 μL的蛋白酶K,混匀;37 ℃温育1 h;加入100 μL 5 mol/L NaCl,充分混匀,再加入80 μL CTAB/NaCl溶液,混匀;65 ℃温育10 min;加入等体积、体积比24∶1的氯仿∶异戊醇溶液,12000 r/min离心4~5 min;将上清液转入一只新管中,加入0.6~0.8倍体积的异丙醇,轻轻混合直到DNA沉淀下来,12000 r/min离心10 min;加入1 mL 70%乙醇洗涤,12000 r/min离心10 min,弃乙醇;在洁净工作台中干燥,重溶于50 μL TE缓冲液或者去离子水中。
1.2.4 16S rDNA测序及构建系统发育树 传统的生化方法对微生物进行正确的分类存在较大的困难,而随着分子生物学技术的发展核酸序列分析以及构建系统发育树已被广泛地应用于微生物的鉴定中,通过序列分析,可以快速准确地获得基因序列,再通过系统发育树的构建从而分析出微生物种类。该方法是以纯化菌的DNA为模板,利用DNA条形码技术,采用PCR方法扩增各纯化菌的16S rDNA片段测序,测序后在将DNA序列在GeneBank进行比对(https://cipotato.org/genebankcip/)。引物为27F:5′-AGAGTTTGATCCTGGCTCAG-3′,反向:1492R:5′-TACGG(C/T)TACCTTGTTACGACTT-3′。PCR扩增体系(30 μL):2×PCR Mix15 μL,DNA模板1 μL,上下游引物各1 μL,无菌水12 μL。PCR扩增程序[10]:95 ℃预变性5 min;95 ℃变性30 s;58 ℃退火30 s;72 ℃延伸45 s;35个循环;72 ℃末端延伸5 min;4 ℃保温。将PCR扩增产物送至天一辉远生物科技有限公司进行基因测序。将所得目的序列使用NCBI的BLAST程序对获得的16S rDNA序列进行同源序列比对,合理选取相似性高的序列,最后使用MEGA7.0.26软件(美国MEGA公司)的邻接法(Neighbor joining)构建系统发育树[17]。
2 结果与分析
2.1 细菌分离纯化
从鮰鱼背部取肉分离纯化得到3株菌株,编号为x1-1、y1-1、y2-1。其中:菌株x1-1在血液琼脂培养基上为圆形、湿润、灰白色菌落,取单菌落革兰氏镜检为革兰氏阴性短杆菌(图1A);菌株y1-1在营养琼脂培养基上为圆形、光滑湿润、乳白色菌落,取单菌落革兰氏镜检为革兰氏阴性短杆菌(图1B);菌株y2-1在营养琼脂培养基上为圆形、扁平湿润、乳白色菌落,取单菌落革兰氏镜检为革兰氏阴性短杆菌(图1C)。
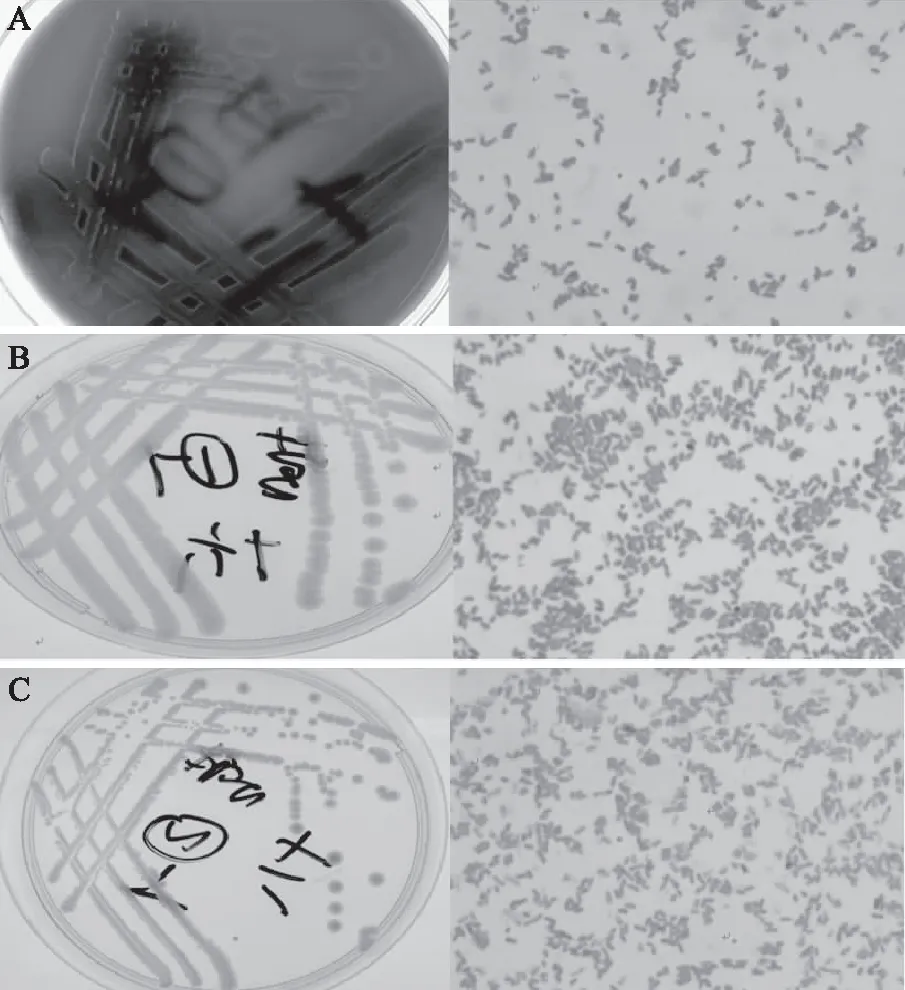
图1 三种腐败菌的菌落特征、形态特征
2.2 生化鉴定
将纯化的菌株接种至生化微量鉴定管中在37 ℃培养24 h,观测其结果为:x1-1硝酸盐还原、乙酰胺、非木糖、西蒙氏柠檬酸盐、DNA、OF为阴性,精氨酸、非麦芽糖、非甘露糖、氧化酶为阳性;y1-1硝酸盐还原、非木糖、西蒙氏柠檬酸盐、DNA、OF为阴性,精氨酸、乙酰胺、非麦芽糖、非甘露糖、氧化酶为阳性;y2-1乙酰胺、非木糖、非甘露糖、西蒙氏柠檬酸盐、DNA、OF为阴性,硝酸盐还原、精氨酸、非麦芽糖、氧化酶为阳性。与《常见细菌系统鉴定手册》[15]比对表明:x1-1与嗜水气单胞菌的生化特性基本一致;y1-1与维氏气单胞菌的生化特性基本一致;y2-1异嗜糖气单胞菌的生化特性基本一致。结果见表1。

表1 纯化菌种生化鉴定结果
2.3 16S rDNA基因测序及系统发育树分析
分别以菌株x1-1、y1-1、y2-1的DNA为模板,通过PCR扩增细菌16S rDNA片段。通过电泳检测发现:三个菌株均约在1500 bp处有明显目的条带(图2)。进一步对16S rDNA测序表明:菌株x1-1目的片段大小为1398 bp;菌株y1-1目的片段大小为1408 bp;菌株y2-1目的片段大小为1409 bp。将各菌落的16S rDNA片段通过Blast检索系统进行序列比对分析以及构建系统进化树,结果表明:x1-1与嗜水气单胞菌(A.hydrophila)亲缘关系最近,达99.9%(图3A);y1-1与维氏气单胞菌(A.veronii)亲缘关系最近达99.79%(图3B);y2-1异嗜糖气单胞菌(A.allosacharophila)亲缘关系最近,达99.8%(图3C)。
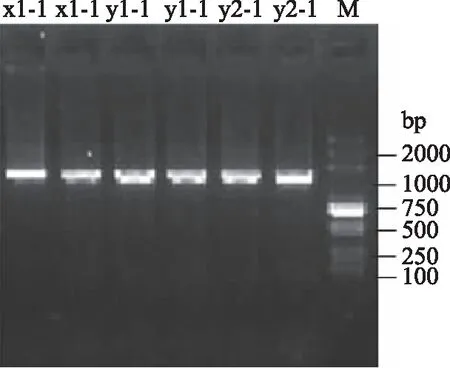
图2 纯化菌种16S rDNA电泳图

图3 三种腐败菌的系统进化树
3 讨论
气单胞菌属(Aeromonasspp.)在自然界中广泛存在,主要来源于水源、土壤以及人的粪便中,可在0~5 ℃生长,因此在水产品冷藏的条件下极易成为优势菌种[18],是水生动物中主要的致病菌之一,能导致多种鱼类感染患病,一旦感染将迅速流行,造成巨大的经济损失[19]。现阶段对气单胞菌属的研究主要集中于水生动物,范围涉及嗜水气单胞菌(A.hydrophila)、维氏气单胞菌(A.veronii)、豚鼠气单胞菌(A.caviae)、温和气单胞菌(A.sobria)等。如蒋慧亮等[20]从4 ℃冷藏的河蚌肉中分离出优势腐败菌是嗜水气单胞菌(A.hydrophila);焦维桢等[21]从真空包装的冷藏鲟鱼中分离得到优势腐败菌为维氏气单胞菌(A.veronii)和路德维希肠杆菌(Enterobacterludwigii),与本实验结果基本一致。但根据最新对冷藏斑点叉尾鮰优势腐败菌的研究[22]表明4 ℃下希瓦氏菌(Shewanallaspp)是鮰鱼4 ℃冷藏中主要的致腐菌群,这可能是由于该实验与本实验在鱼肉的取样环境、储藏时间、储藏环境、纯化程度等因素存在差异导致。
腐败微生物能够分解鱼肉的脂肪和蛋白质等营养物质,积累导致鱼肉产生不愉快气味甚至变色、变味等的胺、吲哚、硫化氢、有机酸、醛和酮等腐败代谢产物[23]。结合其他学者研究[17,23-25]及本实验生化鉴定结果表明:三种气单胞菌均为非发酵型革兰氏阴性菌,均可利用葡萄糖、木糖作为碳源,但不利用麦芽糖、甘露糖、西蒙氏柠酸盐;可利用蛋白质中的精氨酸等作为氮源,从而这三种气单胞菌消耗鱼肉中的蛋白质、糖类等营养物质,使鱼肉变质且自身大量繁殖。根据陈稚峰等[16,26-29]对鱼源、虾源中分离出的气单胞菌的群体感应现象的研究表明:气单胞菌能够分泌高丝氨酸内酯(AHLs),促进蛋白酶的分泌及自身生物膜的形成,进而使食物中的蛋白质降解成小分子肽等,增强自身抗逆性,加速水产品腐败。根据李苗云等[30]对冷却猪肉贮藏过程中分离出的主要腐败菌的聚类分析的研究表明:影响肉品腐败不仅仅是单独的某一类微生物作用的结果,而是多种腐败菌相互作用所导致的。单一的气单胞菌的致腐能力并不强,但在混合细菌(包括气单胞菌、热杀索丝菌、肠杆菌科细菌和假单胞菌)的共同作用下,肉品腐败速度加快,并且在导致产品腐败的进程中,气单胞菌对腐败的群体效应起关键作用。即气单胞菌可能与其他腐败菌共同腐败,但这些仍需要进一步研究。
4 结论
综上所述,本实验结合细菌形态观察、生化鉴定、16S rDNA序列测定等方法共同确定冷藏鮰鱼片优势腐败菌的种类,结果真实可靠。研究表明:在冷藏条件下,鮰鱼片的优势腐败菌主要为气单胞菌属中的嗜水气单胞菌(Aeromonashydrophila)、维氏气单胞菌(Aeromonasveronii)、异嗜糖气单胞菌(Aeromonasallosaccharophila)。
研究真空包装的冷藏鮰鱼片中的优势腐败菌,不仅能够了解真空包装以及低温条件下对腐败菌生长情况的影响,还能了解真空包装以及低温条件下仍能导致鮰鱼肉质腐败的腐败菌种类,可为鮰鱼在商品流通过程中肉质腐败的防控提供研究方向。
