牛磺酸对肉鸡生长性能、肉品质及肌肉抗氧化功能的影响
2020-09-21韩红丽张婧菲沈明明何家乐杨培萱
韩红丽,张婧菲,沈明明,何家乐,刘 佳,杨培萱,王 恬
(南京农业大学动物科技学院,江苏 南京 210095)
牛磺酸(taurine,TAU)是一种含硫的β氨基酸,以高浓度存在于绝大部分动物细胞中[1]。在1827年,研究者首次从牛胆汁中分离出TAU[2]。TAU已广泛应用于动物生产中,具有促进动物生长、提高机体抗氧化能力和免疫力等多种生物学功能。日粮添加适量TAU能改善肉鸡的生长性能、免疫功能、总抗氧化能力(total antioxidant capacity,T-AOC)和脂质代谢[3]。饲料中添加适宜剂量的TAU,能提高鲤鱼肠道超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)和谷胱甘肽过氧化物酶(glutathione peroxidase,GPx)的活性,抑制丙二醛(malondiadehyde,MDA)的生成,增强鲤鱼的抗氧化能力[4]。此外,TAU的前体物质(半胱氨酸、半胱胺、亚牛磺酸等)也具有抗氧化活性,可有效清除活性氧自由基,抑制脂质过氧化反应[5]。
近几十年来,肉鸡产业发展迅速,全球肉鸡消费量仅次于猪肉,但是由于肉鸡肌肉中富含多不饱和脂肪酸,易氧化酸败,引发肌肉变质。现代养殖环境中的各种应激(如热应激、免疫应激、屠宰应激等)均会加速机体脂质氧化,引起肌肉蛋白质和脂质过氧化,影响肌肉的肉品质(如肉品失色、滴水损失增加等),从而对肉品质产生不良影响。氧化应激通过改变蛋白质结合氢和毛细管结合水分子的能力,影响肌肉的保水性[6]。因此,通过提高肌肉的抗氧化能力,能有效改善肌肉品质。然而,目前关于TAU在肉鸡方面的研究多集中于从抗氧化酶活性的角度来探讨TAU的抗氧化作用,但有关TAU对肉鸡肌肉抗氧化功能及相关基因mRNA表达水平对肌肉品质的影响研究仍十分缺乏。因此,本实验旨在研究日粮中添加不同水平的TAU对肉鸡生长性能、肌肉品质、胸肌抗氧化功能的影响,探讨肌肉抗氧化与肌肉品质之间的关系,同时筛选出TAU在肉鸡日粮中的适宜添加水平,为生产中以营养调控的方式改善肉鸡的肌肉品质提供新的思路和方法,同时也为TAU在提高肉品质健康方面的研究提供参考。
1 材料与方法
1.1 材料与试剂
TAU(纯度>98.5%)购自江阴华昌食品添加剂有限公司;‘艾拔益加’(Arbor Acres,AA)肉鸡购自安徽和威集团;DEPC水购自上海碧云天生物技术有限公司。T-AOC、GPx、SOD、CAT和MDA试剂盒均购自南京建成生物工程研究所;TRIzol试剂、反转录试剂盒、聚合酶链式反应(polymerase chain reaction,PCR)试剂盒均购自宝生物工程(大连)有限公司。
1.2 仪器与设备
Nanodrop ND-2000c分光光度计、Muliskan Go全波长酶标仪、Q u a n t S t u d i o 5 荧光定量P C R 仪美 国T h e r m o S c i e n t i f i c 公 司;C R-4 0 0 型 色 度仪日 本K o n i c a M i n o l t a 公 司;H I 9 0 2 5 型p H 计意大利Hanna公司;C-LM3B嫩度仪 东北农业大学;5804R离心机 德国Eppendorf公司;Secura125-1CN电子天平 赛多利斯科学仪器(北京)有限公司。
1.3 方法
1.3.1 实验分组及设计
实验采用随机单因素试验进行分组设计,选择1 日龄初始体质量相近的健康AA肉鸡384 只,随机分为4 组,每组12 个重复,每个重复8 只肉鸡。对照组(CON组)肉鸡饲喂玉米-豆粕型基础日粮,实验组肉鸡分别在基础日粮的基础上添加0.25%(TAU1组)、0.5%(TAU2组)、0.75%(TAU3组)的TAU。实验周期为42 d,分为前期(1~21 d)和后期(22~42 d)。实验肉鸡采用3 层叠笼饲养。在实验期间,自由采食和饮水,每日清粪,保持舍内的通风换气。实验过程中,严格按照常规程序进行免疫处理。基础日粮参考National Research Council(1994)《肉鸡饲养标准》完成配制[3],配方及营养水平见表1。

表 1 日粮配方及营养水平Table 1 Ingredients and nutrient contents of experimental diets used in this study
1.3.2 样品采集
于实验第42天,从每个重复中随机选1 只体质量接近于该重复平均体质量的肉鸡,此前禁食12 h。于颈动脉处放血,待其死亡后立即解剖,在每只肉鸡的同一部位取适量胸肌样品,迅速置于-80 ℃冰箱保存备用,同时取另一侧胸肌用于测定肉品质。
1.3.3 指标测定
1.3.3.1 生长性能测定
于实验第21天和第42天分别禁食12 h后空腹称体质量,以每个重复为单位记录采食量,并以此为基础计算平均日采食量、平均日增体质量和料重比(平均日采食量与平均日增体质量之比)[3]。
1.3.3.2 肉品质测定
pH值测定:于实验第42天肉鸡屠宰后,迅速取下一侧胸肌置于冰上,45 min时测定1 次肉样的pH值,然后置于4 ℃冰箱,24 h后再次测定肉样的pH值。每个样品于不同位置测量3 次,结果取其平均值[7]。
肉色测定:胸肌样品在4 ℃存放24 h后取出,使用色度仪测定样品的明度(L*)、红度(a*)和黄度(b*),每个样品于3 个不同位置进行测定,结果取其平均值[8]。
滴水损失率测定:于实验第42天,肉鸡屠宰后,将肌肉修整为3 cm×2 cm×1 cm的肉块,称质量(m1/g),然后用细铁丝钩住肉样,使肌纤维垂直向下,悬吊于4 ℃冰箱,24 h后取出肉样后称质量(m2/g),按式(1)计算24 h滴水损失率,再过24 h后取出肉样称质量(m3/g),按式(2)计算48 h滴水损失率[9]。
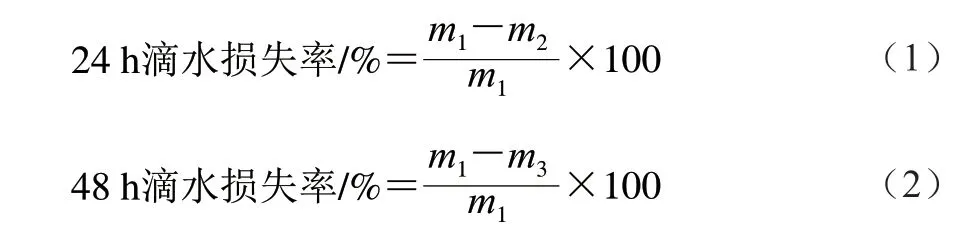
蒸煮损失率测定:于实验第42天肉鸡屠宰后,用取样器切取肉块,去除结缔组织,用塑料袋包装,在0~4 ℃冰箱熟化24 h,称质量(m1/g)。把肌肉放到水浴锅中加热,加热到肉中心温度为70 ℃,水浴温度在75~80 ℃,取出肉样冷却后称质量(m2/g)[9]。按式(3)计算蒸煮损失率。

剪切力测定:将测定蒸煮损失率后的样品顺着肌纤维方向切为截面为1 cm×1 cm的3 个肉条,再以垂直肌纤维方向用嫩度仪对其进行剪切,每个肉条剪切1 次,结果取3 次剪切力读数的平均值[8]。
1.3.3.3 肌肉抗氧化能力测定
称取约0.5 g的肌肉样品,加入4.5 mL生理盐水(质量分数0.75%的NaCl溶液),在冰浴条件下充分匀浆,然后4 ℃条件下4 000×g离心10 min,吸取分装上清液,置于-20 ℃条件下储存待测[10]。测定肌肉(1 mg蛋白中)T-AOC、GPx活力、SOD活力、CAT活力和MDA含量。
1.3.3.4 肌肉抗氧化相关基因mRNA的表达测定
总RNA的提取:提前将所用试剂预冷,取出-80 ℃条件下保存的肌肉样品,切取约50 mg肌肉,采用TRIzol试剂从肌肉中提取总RNA,使用分光光度计测定OD260nm和OD280nm,通过观察两者的比值即OD260nm/OD280nm来评估总RNA的浓度和纯度,比值介于1.8~2.1表示RNA的纯度较高、未被污染。
RNA反转录:将不同样品的RNA浓度用DEPC水统一稀释成500 ng/μL,利用反转录试剂盒合成cDNA。反转录的反应条件为:37 ℃下反应15 min,85 ℃下变形15 s,最后降温至4 ℃。
荧光定量PCR:利用实时荧光定量PCR试剂盒进行。PCR程序为:95 ℃、30 s,95 ℃、5 s,60 ℃、31 s,40 个循环。目的基因的相对表达量以β-Actin作为内参基因,采用比较2-ΔΔCt法进行计算。所有基因序列均从GenBank中获取,引物序列见表2,由生工生物工程(上海)股份有限公司合成。

表 2 目的基因及内参基因引物信息Table 2 Primer sequences information for target and reference genes
1.4 数据处理与分析
实验数据采用Excel 2010软件进行初步汇总整理后,采用SPSS Statistics 22软件进行统计,采用单因素方差分析进行差异显著性检验,并采用Duncan’s法进行多重比较,结果用平均值±标准误差表示,当P<0.05时表示差异显著。
2 结果与分析
2.1 添加不同水平的TAU对肉鸡生长性能的影响

表 3 添加不同水平TAU对肉鸡生长性能的影响Table 3 Effects of dietary TAU supplementation at different levels on growth performance of broilers
由表3可知,实验前期,与CON组相比,各TAU处理组均能显著提高肉鸡的平均日增体质量(P<0.05)。其中,TAU1和TAU2组平均日采食量较CON组分别显著提高了8.90%和17.77%。实验后期,与CON组相比,各TAU处理组均能显著提高肉鸡的平均日采食量(P<0.05),其中TAU2组平均日采食量较CON组相比提高了9.94%。从实验全期来看,与CON组相比,各TAU处理组均能显著提高肉鸡的平均日增体质量和平均日采食量(P<0.05)。其中,TAU1和TAU2组的料重比相较CON组分别显著降低了5.33%和1.78%(P<0.05)。
2.2 添加不同水平的TAU对肉鸡胸肌肉品质的影响

表 4 添加不同水平的TAU对肉鸡胸肌肉品质的影响Table 4 Effects of dietary TAU supplementation at different levels on breast muscle meat quality of broilers
由表4可知,与CON组相比,TAU2组胸肌的滴水损失率和蒸煮损失率显著降低(P<0.05)。各TAU处理组pH24h均显著高于CON组(P<0.05),但TAU处理组之间差异不显著(P>0.05)。就胸肌的b*值而言,TAU1和TAU2组显著低于CON组(P<0.05)。各组间的胸肌剪切力和L*值差异均不显著(P>0.05)。
2.3 添加不同水平的TAU对肉鸡胸肌肉抗氧化指标的影响

表 5 添加不同水平的TAU对肉鸡胸肌肉抗氧化指标的影响Table 5 Effects of dietary TAU supplementation at different levels on the antioxidant capacity of breast muscle in broilers
由表5可知,与CON组相比,各TAU处理组均能显著提高肉鸡胸肌中GPx活力(P<0.05),降低MDA含量(P<0.05)。TAU1组SOD活力显著高于CON组(P<0.05)。
2.4 添加不同水平的TAU对肉鸡胸肌抗氧化相关基因mRNA相对表达量的影响

图 1 TAU对肉鸡胸肌抗氧化相关基因mRNA相对表达量的影响Fig. 1 Effects of dietary TAU supplementation at different levels on antioxidant enzyme mRNA expression in broilers
由图1可知,与CON组相比,各TAU处理组均能显著提高肉鸡胸肌中GPx4、GPx及SOD1mRNA相对表达量(P<0.05),但TAU1、TAU2和TAU3各处理组之间差异不显著(P>0.05)。与CON组相比,各TAU处理组均能显著降低CAT的相对表达量(P<0.05)。
3 讨 论
3.1 TAU添加水平对肉鸡生长性能的影响
目前,关于TAU对动物生长性能的影响已有一定的研究。李丽娟等[10]的研究结果表明,在生长前期日粮中添加TAU可不同程度地提高肉雏鸡的平均日增体质量,降低料重比。黄春喜等[11]研究发现,日粮添加TAU可以显著降低肉仔鸡早期(1~7 d)的料重比。Liu Yue等[12]发现饲粮添加TAU能提高仔猪的生长性能,促进肠道发育。本实验结果表明,从全期来看,日粮添加TAU能显著提高肉鸡的平均日增体质量和平均日采食量,其中添加0.25%和0.5%的TAU能显著降低肉鸡的料重比,这与上述研究结果基本一致。TAU的促生长作用可能与它的抗氧化作用有关,TAU可缓解活性氧对机体脂质的氧化损伤,提高动物体对脂肪的利用,进而促进动物的生长。也有研究表明,在植物蛋白的饲粮中添加TAU对斑马鱼的生长性能没有显著影响[13]。因此推测,造成这种差异的原因可能与日粮类型、物种、生长阶段、饲养环境和TAU的添加量等因素有关。
3.2 TAU添加水平对肉鸡肉品质的影响
影响肌肉品质的因素有很多,包括pH值、系水力、剪切力和肉色等。剪切力是评价肌肉嫩度的重要指标,剪切力越小,说明肌肉的嫩度越好[14]。另外,肌原纤维的状态、肌肉中结缔组织的含量、结缔组织中胶原蛋白的种类和交联程度均是影响肉嫩度的重要因素[15]。本实验结果表明,在日粮中添加适量的TAU能降低肌肉的剪切力、提高肌肉的嫩度、降低肌肉的滴水损失,其中以0.25%和0.5%的添加量效果最佳,这与臧素敏等[16]的研究结果一致。钙蛋白酶的氧化可以抑制蛋白质降解,而相关研究表明,动物体内抗氧化防御系统的提高会影响钙蛋白酶活性、蛋白质水解,从而改善肌肉的系水力和嫩度[17],表明TAU的添加可能是通过上述途径提高肌肉的系水力和嫩度。
pH值是影响肌肉系水力的重要因素,肌肉pH值的升高在一定程度上会减少蛋白质的变性,使肌原纤维蛋白质偏离其等电点,减弱蛋白质之间的相互作用,进而降低滴水损失和蒸煮损失,提高肌肉的系水力[9]。此外,pH值的变化能够在一定程度上反映肉鸡屠宰后机体糖原酵解的速度。肉鸡屠宰后,肌肉中乳酸菌对糖原进行无氧酵解从而产生乳酸,乳酸积累会导致肌肉酸度增加,pH值下降[18]。本实验发现,肉鸡屠宰后pH45min和pH24h实验组均高于CON组,其中以0.5%的TAU添加量效果最佳。魏源等[19]的研究结果表明,服用TAU能降低运动大鼠乳酸含量,由此推测TAU可能通过减少肌肉中乳酸的积累来提高pH值,延缓肌肉变质的速度,改善肉鸡的肌肉品质。但Huang Chunxi等[20]研究结果表明日粮中添加不同水平的TAU对肌肉pH值和肉色均未产生显著影响,这种差异的原因可能与物种、饲料组成以及添加剂量等因素有关。
肉色是肌肉外观评定的重要指标,是肌肉的生理学、生物化学和微生物学变化的外部体现[21]。实验结果显示,在日粮中添加TAU能显著降低肉鸡胸肌的b*值,b*值表示肉样的黄度,b*值越低肉色越好。曾得寿等[22]发现在肉仔鸡日粮中添加0.2%TAU可以显著提高肉鸡胸肌的肉色。肉的色泽主要受肌红蛋白含量的影响,肌红蛋白含量越多,肉色越深。研究表明肌肉组织内的氧化会导致高铁肌红蛋白的生成,使得肉的表观色泽变差,其机制涉及不饱和脂肪酸的过氧化,而抗氧化剂的加入可以保护脂质不被氧化,并稳定氧肌红蛋白的含量[23]。由此推测添加适量的TAU可能通过其抗氧化作用抑制脂质过氧化,在一定程度上稳定氧肌红蛋白的含量,进而改善肌肉胸肌的肉色[24]。
3.3 TAU添加水平对肉鸡肌肉抗氧化功能的影响
TAU具有多种生物学功能,其中抗氧化功能是TAU最为重要的生物学功能之一。肉品质与抗氧化属性密切相关。脂质过氧化是导致肉品质恶化的重要原因之一,它可以降低肌肉的营养价值、影响风味和质地、改变肌肉的外观,因此提高肌肉的抗氧化功能,对于改善肌肉品质具有重要的意义。MDA是脂质过氧化代谢的代表性产物之一,其含量的高低直接反映脂质过氧化损伤的程度。在正常生理状态下,体内的抗氧化酶(如GPx、SOD)可发挥协同作用,中和超氧化物阴离子自由基和过氧化氢,减轻氧自由基链式反应对机体的损害[25]。相关研究已经证明,在日粮中添加TAU能通过提高机体抗氧化酶的活性,降低MDA的含量,从而提高机体的抗氧化功能[26-27]。本实验结果表明,在日粮中添加TAU能提高GPx和SOD活力,降低MDA的含量,与上述研究结果一致。研究表明,抑制肌肉的脂质过氧化,可减弱蛋白质降解,降低肌原纤维间的水分储备,从而降低肌肉的系水力[28]。实验所观察到的胸肌肌肉蒸煮损失的减少可能是因为肌肉的系水力与蛋白质水解存在一定的关系[29],TAU的添加降低了肌肉蛋白质和脂质的过氧化,进而提高肌肉的保水性。GPx4是机体内酶类抗氧化系统中的重要成员,主要作用是清除膜或脂蛋白中的脂质过氧化物。有研究表明,细胞膜的完整性与滴水损失有关,由于肌肉细胞膜中含有大量不饱和脂肪酸和磷脂,易受自由基的攻击而诱发脂质过氧化,从而破坏细胞膜的结构,增加鸡肉水分的渗出。由此推测,TAU提高肌肉系水力可能是通过上调肌肉中GPx4基因的表达,降低肌肉中MDA的含量来实现的[30]。同时,使蒸煮损失减少,细胞失水皱缩减少,从而改善肌肉的色泽。此外,机体内脂质过氧化产物含量的减少会减缓氧合肌红蛋白转化为高铁肌红蛋白,进而改善肌肉颜色[31],最终缓解肌肉氧化变质,提高肌肉嫩度,但具体机制有待进一步研究。本实验发现TAU可以上调肌肉中GPx和SOD1基因的表达,这个结果表明TAU可能通过调控抗氧化酶的基因表达,从分子水平来提高肉鸡胸肌的抗氧化能力,这也与TAU能提高GPx和SOD活力结果一致。
综上,在日粮中添加TAU可以提高肉鸡的生长性能、改善肉鸡胸肌的肉品质和抗氧化功能。添加TAU可能与其抑制脂质过氧化、提高胸肌抗氧化基因mRNA的表达水平有关。本实验条件下,肉鸡日粮中TAU添加量以0.5%效果最佳,表明TAU对于改善肉品质健康具有重要的作用,为其在食品科学领域的研究提供了参考。
