西蓝花废弃茎叶固态发酵饲料的营养成分及活性代谢产物的分析研究
2020-09-18姚晓红汤江武吴逸飞李园成
孙 宏,姚晓红,汤江武,吴逸飞,王 新,李园成,沈 琦
(植物保护与微生物研究所,浙江省农业科学院,浙江杭州 310021)
西蓝花(Brassica oleraceaL.var.italica)又名绿菜花,其蛋白质、维生素和矿物质含量高,富含酚类化合物,可有效预防人体心脑血管疾病和癌症的发生。目前,中国西蓝花种植面积已超过2.7 万hm2,年产量超过100万t[1-2],但新鲜西蓝花在采摘过程中会产生大量的废弃茎叶,其含水率在90% 以上,极易腐烂,导致臭气污染和附近水源地污染[2],严重威胁环境安全。国内外研究尝试通过制备青贮饲料、生物发酵饲料和西蓝花茎叶蛋白粉等方式来实现西蓝花废弃茎叶的饲料化利用,取得了较好的效果[3-5]。由于将西蓝花废弃茎叶制备为发酵饲料具有成本较低、工艺控制程度高、发酵周期短等优势,较适宜于规模化推广应用[5-6]。但目前该方面的研究仍较少,且多集中于动物应用效果评价,缺乏对发酵前后西蓝花茎叶中营养成分,特别是活性成分的全面分析评估。生物发酵饲料是一个复合体系,包括发酵底物原料及其降解产物、微生物生长代谢产物等多种复杂的小分子化合物,这也增加了其全面分析的难度。
代谢组学技术(Metabolomics)是一种基于组学思想和高分辨率色谱、质谱分析手段,用于研究生物代谢途径及代谢产物的技术。自20 世纪末以来,代谢组学技术已逐渐应用于动物血液、食品发酵乳制品、中草药等复杂代谢物的定性和定量研究[7]。但目前其在固态发酵过程中的应用报道仍较少[8]。本试验尝试采用代谢组学技术对西蓝花茎叶与玉米皮、米糠和麸皮等原料混合后固态发酵制备的发酵饲料中活性产物进行分析,探讨固态发酵对混合发酵原料中代谢物的影响,为完善和评价西蓝花茎叶制备发酵饲料的实际效果提供基础资料。
1 材料与方法
1.1 西蓝花原料及发酵菌株 西蓝花茎叶、玉米皮、米糠和麸皮均购自浙江台州天莱生物科技有限公司。短小芽孢杆菌(Bacillus pumilus)XLJ2-1、酿酒酵母(Saccharomyces cerevisiae)99-1 和干酪乳杆菌(Lactobacillus casei)Lvk1 均为本实验室培养选育和保藏。新鲜西蓝花茎叶原料营养成分(鲜料)实测值分别为粗蛋白质2.62%、水分85.0%、粗纤维4.8%、粗脂肪0.5%、粗灰分1.6%。
1.2 固态发酵西蓝花的制备 发酵原料为含有50%(w/w)西蓝花茎叶碎渣、20%玉米皮、20%米糠和10%麸皮的混合原料。将上述发酵底物混合均匀后,接种1.0%(v/w)含有短小芽孢杆菌XLJ2-1、酿酒酵母99-1 和干酪乳杆菌Lvk1 的混合菌液(比例为1:1:1,v/v;混合菌液中初始活菌数分别为2.6×108、0.8×108、2.2×108CFU/mL)。取2.5 kg 西蓝花废弃茎叶混合原料装入饲料厌氧呼吸发酵袋中,于30℃下恒温培养15 d。以接种前的混合原料为对照,每组设置5 个重复。
1.3 检测指标与方法
1.3.1 常规营养成分分析及活菌数测定 取接种前与发酵后的新鲜西蓝花茎叶样品200 g,参照文献[9]的报道,测定西蓝花样品的粗蛋白质、粗脂肪、粗纤维、水分和灰分等常规营养成分。参照《食品微生物学检验-菌落总数测定》(GB 4789.2-2016)检测西蓝花发酵后的芽孢杆菌、乳酸菌和酵母菌数量。
1.3.2 发酵样品的代谢组测定
1.3.2.1 样品提取及预处理 取发酵前后新鲜西蓝花样品1 g,加入50 mL 液氮研磨均匀,取0.1 g 研磨样品放入无菌1.5 mL 离心管中,加入0.4 mL 预冷的提取液(甲醇:乙腈 :水,4:4:2,v/v),涡旋混合1 min 后,–20℃下静置60 min,14 000 r/min 离心20 min,取离心上清真空干燥至干。质谱分析前加入100 μL 浓度为50%的乙腈水溶液(乙腈:水,1:1,v/v)复溶。将复溶后的样品经过0.22 μm 微孔滤膜过滤后,置于液相样品瓶中,待测。
1.3.2.2 UPLC-Q-TOF 检测 样品采用UPLC 系统(Agilent 1290,CA,USA)进行色谱分离,其中色谱柱采用C18反向色谱柱(1.7 μm,2.1×100 mm),柱温25℃、流速0.3 mL/min、进样体积2 μL。流动相A 为含有终浓度25 mmol/L 乙酸铵和25 mmol/L 氨水的水溶液,流动相B 为乙腈。梯度洗脱程序:0~1 min,95% 流动相B;1~14 min,流动相B 浓度从95%下降到65%;14~16 min,流动相B 从65%继续下降到40%;16~18 min 维持40%流动相B 浓度;18~20 min,95%流动相B 冲洗。
样品经UPLC 分离后采用AB SCIEX TOF 6600 质谱仪进行分析。电喷雾离子源(ESI);四级杆飞行时间串联质量分析器;扫描采用正负离子两种模式进行,离子源温度600℃,毛细管电压2.0 kV,质核比扫描范围60~1 000 m/z。
1.3.2.3 差异代谢物鉴定及代谢途径富集分析 将总离子流图谱转换为格式文件mzXML,然后利用XCMS 程序提取原始数据,并进行峰对齐、保留时间矫正和提取峰积分面积,根据保留时间、精准质量数匹配原则(误差小于20 mg/kg)和二级图谱匹配方式等,与人类代谢组数据库(HMDB)、微生物代谢组数据库(YMDB、HMDB)和自建数据库中相关信息进行匹配,从而完成代谢物鉴定[10]。对XCMS 输出的保留时间-质核比数据矩阵,采用SIMCA-P11 软件进行多维统计(PCA)分析区分发酵前后西蓝花茎叶样品。以差异倍数分析Ratio>2 或<0.5 为筛选标准,找出西蓝花茎叶发酵前后的差异代谢物;以P<0.05 为标准,将候选差异代谢物的相对含量进一步进行t检验验证。最后,为了阐明差异代谢物的生物学功能,使用MetaboAnalyst 3.0 软件结合KEGG 等数据库,通过富集倍数检验来确定受到显著影响的代谢途径[11]。
1.3.3 活性产物分析验证 发酵西蓝花样品中粗多糖的测定参照《植物源食品中粗多糖的测定》(SN/T 4260-2015)进行;酸溶蛋白含量参照国标《大豆肽粉》(GB/T 22492-2008)的测定方法进行;总酸参照《食品中总酸的测定方法》(GB/T 12456-2008)测定;γ-氨基丁酸的测定参照顾宇翔等[12]的报道进行。
1.4 统计分析 利用SPSS 16.0 软件进行统计分析,两组间差异分析采用t检验。所有数据以平均值± 标准差表示,P<0.05 表示差异显著。
2 结果与分析
2.1 西蓝花茎叶混合原料发酵前后常规营养成分分析及活菌数测定 西蓝花茎叶混合原料样品经过15 d 固态发酵后呈黄色,有较强的酸香气味。由表1 可知,发酵后西蓝花茎叶混合原料中芽孢杆菌、乳酸菌和酵母菌分别为3.5×106、7.6×106、0.2×106CFU/g。与发酵前相比,发酵后的西蓝花茎叶混合原料中粗蛋白质、粗脂肪、粗纤维、粗灰分和水分的含量均无显著差异。
2.2 西蓝花茎叶混合原料发酵前后代谢物轮廓分析 图1 为西蓝花茎叶混合原料发酵前后代谢物的PCA 得分图。正、负离子检测模式下模型解释率(R2X)分别为0.668 和0.686,满足数据拟合要求。同时,发酵前后样品获得了有效分离。
2.3 发酵前后西蓝花茎叶混合原料中差异代谢物的鉴定由表2 可知,西蓝花茎叶混合原料发酵前后的显著差异代谢物共有35 种,主要包括脂肪酸(有机酸)类、多糖及其衍生物类、核酸类、氨基酸(肽)类、维生素类及其他小分子活性物质。在脂肪酸(有机酸)方面,柠檬酸和异柠檬酸在发酵后降低(P<0.05),D-乳酸、亚油酸、α-亚麻酸含量提高(P<0.05);糖类物质方面,2-酮基-葡萄糖酸和半乳糖酸的含量在发酵后增高(P<0.05),异鼠李糖和甘露庚糖醇含量降低(P<0.05);核酸方面,黄嘌呤、5-甲硫腺苷和腺嘌呤核苷含量均下降(P<0.05);氨基酸(肽)类方面,除谷氨酰-丙氨酸、脯氨酸-苯丙氨酸、甘氨酸-谷氨酸和亮氨酸-丙氨酸含量在发酵后显著降低外,其余11 种肽类物质和组氨酸、L-甲硫氨酸、N-乙酰-谷氨酸的含量均在发酵后显著提高;维生素方面,L-抗坏血酸和硫胺素含量在发酵后提高(P<0.05)。此外,4-吡哆酸、三乙醇胺和薰衣草醇含量在经过固态发酵后降低(P<0.05),而γ-氨基丁酸含量提高(P<0.05)。

表1 发酵前后西蓝花茎叶饲料中营养成分和活菌数含量

图1 发酵西蓝花茎叶样品的PCA 分析
代谢途径富集分析也表明,固态发酵对代谢途径的影响主要集中在蛋白质降解、氨基酸合成、植物次生代谢以及维生素降解途径等方面(图2)。
2.4 发酵西蓝花茎叶混合原料的活性产物分析 由表3可知,发酵西蓝花样品中总酸、酸溶蛋白和γ-氨基丁酸含量均高于发酵前(P<0.05);而发酵前后粗多糖含量无差异。
3 讨 论
3.1 发酵西蓝花茎叶混合原料常规营养成分分析 本研究中西蓝花茎叶原料发酵后颜色加深至黄色,可能与其发酵过程中叶绿素逐渐被破坏有关[13]。田久东等[14]报道西蓝花渣经过3 d 固态发酵后,粗蛋白质含量提高15%,还原糖含量提高50%。但本研究中常规饲料成分在发酵前后无显著差异,这可能与所用的菌种和发酵周期差异有关。本研究采用厌氧呼吸袋作为发酵介质,可制造较好的厌氧环境,也导致了发酵后营养成分与传统固态发酵存在差异[15]。本研究中接种前微生物数量较低可能与西蓝花废弃茎叶在处理过程中采用洗净等加工工艺有关。乳酸菌和芽孢杆菌数量在发酵后仍较高,与前人结果一致[16];而发酵后酵母菌数量较发酵初始有所降低,可能与其在较低pH 下生长受到抑制有关,从而进一步为乳酸菌的生长提供便利[17]。
3.2 固态发酵对西蓝花茎叶饲料代谢物的影响 本研究中发酵前后样品在PCA 分析中获得了有效分离,说明两组之间代谢存在差异[18]。差异代谢物鉴定进一步证实固态发酵处理可引起包括脂肪酸、多糖、核酸、氨基酸、维生素在内的多种代谢物的显著改变,并与途径富集分析的结果一致。代谢通路富集分析可依据代谢物的相关性对通路进行归属化,因而这些受影响的代谢途径与西蓝花固态发酵关系密切,为进一步研究固态发酵对其代谢物影响提供了线索。目前,利用代谢组技术分析固态发酵中代谢物变化的研究仍较少。孙杰等[8]通过GC-MS 代谢组手段分析了酵母液态、固态发酵过程中代谢物差异,发现蛋白质和氨基酸代谢是酵母生长过程中最为重要的差异代谢途径,该结果与本研究结果类似。王越男等[19]在分析脱脂乳和发酵乳样品的差异代谢物时也证实,共有16 种有机酸、20 种多肽、12 种氨基酸和8 种核酸代谢物含量发生显著变化。程新等[11]研究发现乳酸菌发酵受锰离子影响的主要途径集中在糖及衍生物、脂肪酸及其衍生物和氨基酸三大类。本研究结果提示,糖类、脂肪酸和氨基酸代谢途径可能也是西蓝花茎叶固态发酵中受影响的主要代谢途径。
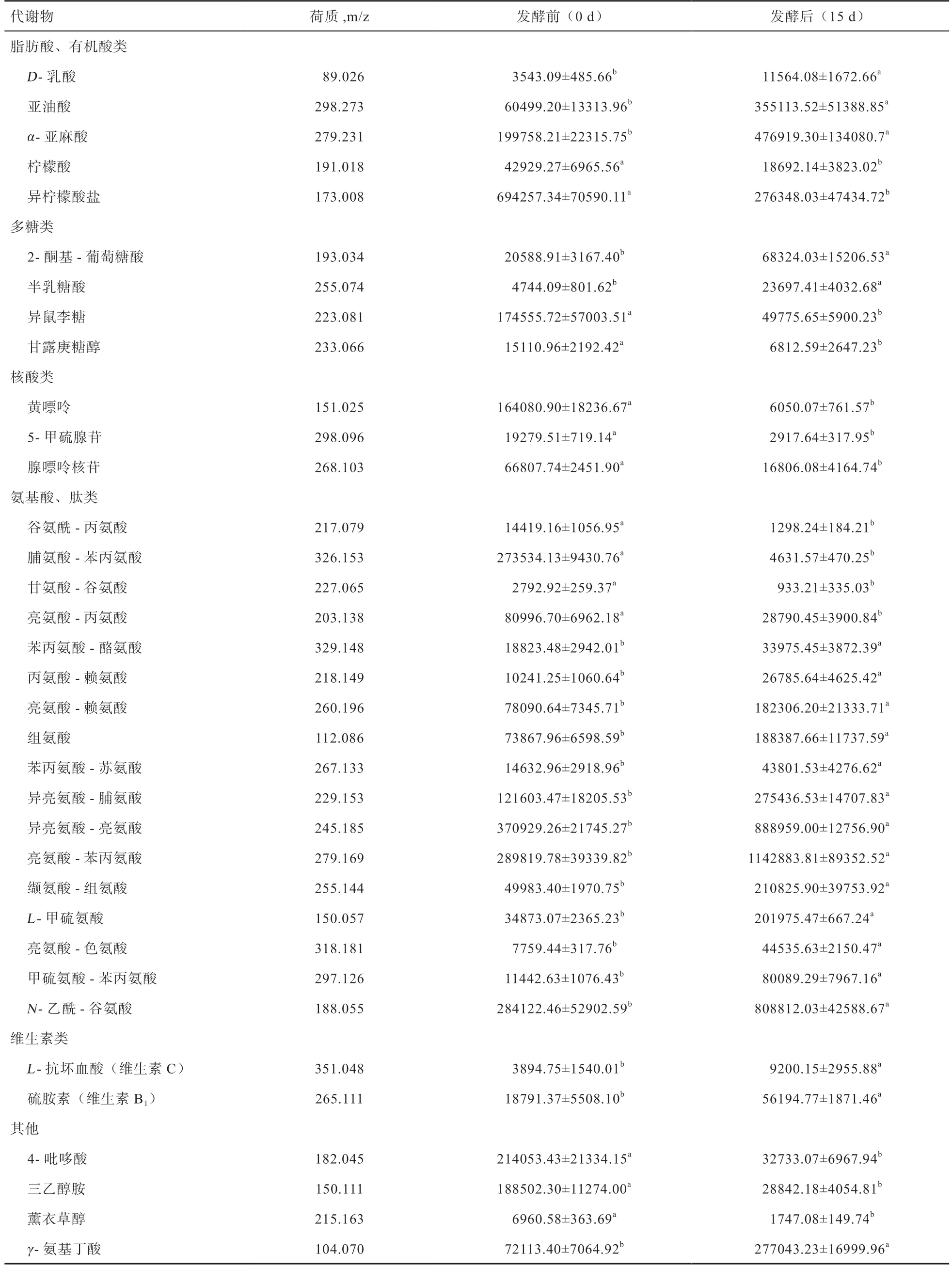
表2 发酵前后西蓝花茎叶混合原料中的差异代谢物及其相对积分含量
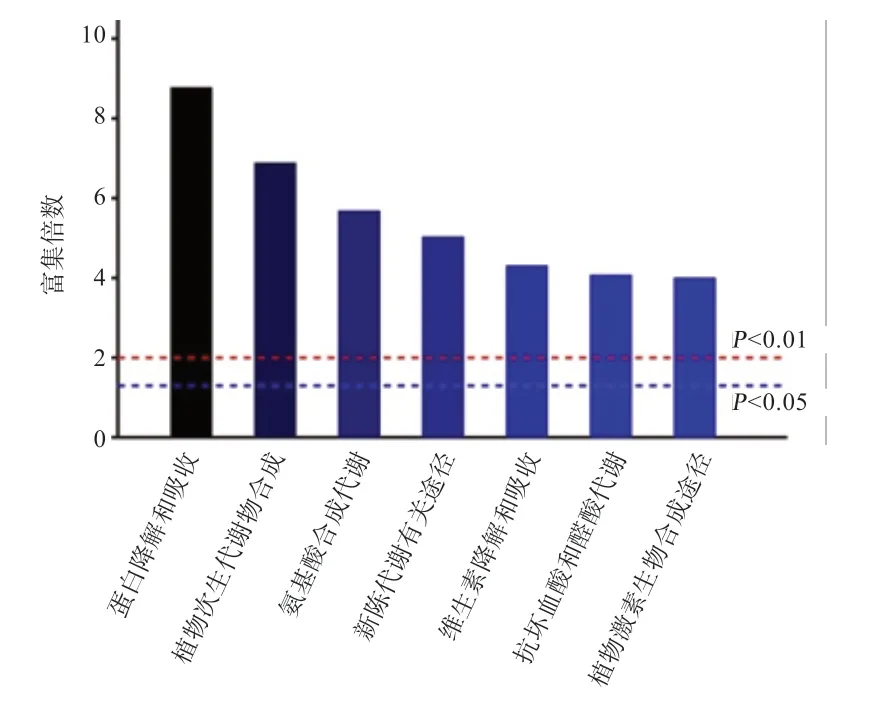
图2 发酵前后西蓝花茎叶混合原料中代谢途径富集分析
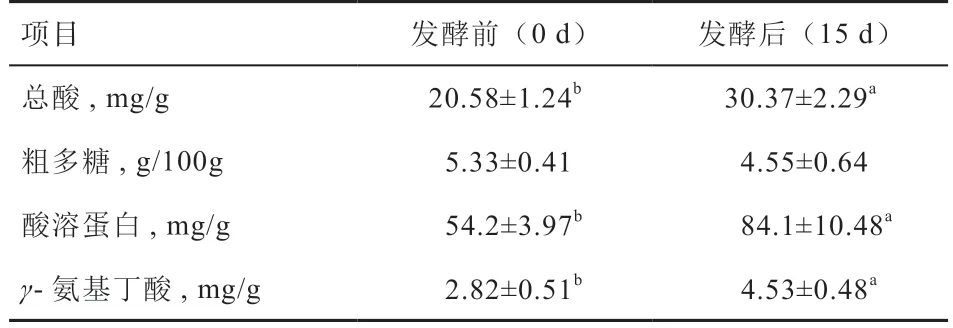
表3 发酵西蓝花茎叶样品中活性代谢产物分析
就具体差异代谢物而言,不同代谢物的变化趋势各不相同。本研究中,代表好氧途径的三羧酸循环中的重要节点化合物(柠檬酸、异柠檬酸)含量在发酵后显著降低,同时乳酸菌重要的代谢产物D-乳酸含量则显著提高。上述代谢物的显著变化表明在本试验发酵条件下,厌氧发酵可能为微生物发酵后期的主要代谢模式。这与本研究发酵周期长达15 d,在发酵中后期,呼吸袋中氧气被芽孢杆菌等好氧微生物耗尽有关[15]。同时,高浓度脂质好氧分解中间产物(亚麻酸、亚油酸)的显著积累也反映发酵后期好氧代谢可能受到了抑制[8],这与Frick 等[20]在酵母发酵中的报道一致。显著提高的2-酮基-葡萄糖酸、半乳糖酸等糖酸类物质可在糖酵解途径中发挥作用,为微生物的厌氧发酵提供能量,这与植物乳杆菌制备发酵乳的代谢方式一致[19]。黄嘌呤、甲硫腺苷和腺嘌呤核苷等核苷物质主要存在于细胞核内,参与胞内的DNA 合成和修复[21]。本研究中发酵后黄嘌呤、5-甲硫腺苷和腺嘌呤核苷含量显著降低,可能与微生物破碎植物细胞分解代谢核苷物质有关。蛋白方面,本试验发酵前后主要差异代谢物为二肽和游离氨基酸,这与在液态发酵乳制品方面的研究结果一致[22]。二肽和游离氨基酸的来源主要为微生物所分泌的蛋白酶对发酵物料的蛋白组分进行分解所致[23]。由于不同微生物对氨基酸的利用存在差异,间接导致了特定短肽和氨基酸含量的升高或降低;而短肽一般较易于被动物肠道吸收利用,且多具有提高动物免疫力、抵抗应激等特定的生物学功能,表明发酵可能提高了西蓝花饲料的营养价值[24]。本研究结果表明,L-抗坏血酸、硫胺素等维生素含量在发酵后显著提高,符合Patel 等[25]所提出的微生物可在发酵过程中参与微生物合成的观点。此外,研究发现γ-氨基丁酸在发酵后会有显著提高[8],表明微生物在次生代谢中参与了该类化合物的合成。范志勇等[26]报道,仔猪日粮中添加γ-氨基丁酸可有效改善猪生长性能,同时显著促进体内生长激素和褪黑素的分泌。本研究结果也显示,发酵后西蓝花茎叶混合原料中γ-氨基丁酸含量显著提高。本试验结果可为进一步解析西蓝花发酵饲料的饲喂效果提供依据。最后,有关4-吡哆酸、三乙醇胺和薰衣草醇等植物激素类前体物质在发酵后显著降低的调控机制仍不明确,推测可能与本研究中涉及到复杂的多菌株联合作用及代谢有关。如聂存喜等[27]曾在利用复合菌协同发酵棉籽粕的代谢组分析中发现了较为明显的菌种发酵互作。后续通过开展单一菌种的发酵并绘制代谢路径图谱将有助于进一步解释该类代谢物差异的原因。需要注意的是,代谢组学的结果更加倾向于对代谢轮廓的描述,本研究虽发现了大量小分子化合物在发酵前后具有显著差异,但仍需进一步验证这些化合物差异。
3.3 发酵西蓝花茎叶饲料的品质分析 一般而言,发酵品质优劣与饲料在发酵后的pH 有密切关系[28]。乳酸菌在发酵过程中可将发酵物料中的糖类转换为乳酸,从而引起pH 和总酸含量的变化[29]。因而发酵西蓝花茎叶中总酸含量的提高与乳酸菌在厌氧发酵后期的大量生长有关[16]。Liu 等[6]报道固态发酵西蓝花茎叶中乙酸、丙酸和丁酸含量分别为5.5、0.033、0.008 g/kg,发酵后pH低于4.5。田久东[14]同样采用玉米皮、麸皮、米糠粕等辅料和西蓝花渣进行固态发酵,发现物料pH 降至4.5 以下,物料中乳酸菌含量可达108 以上。韦仕静等[16]则在制备西蓝花酵素的研究中报道发酵液pH 不断降低至3.51。本研究进一步证实,发酵后西蓝花茎叶饲料中的总酸、酸溶蛋白和γ-氨基丁酸含量显著高于发酵前物料。酸溶蛋白含量是反映发酵后蛋白降解情况的指标,其含量越高表明蛋白分解越多[27]。本研究中酸溶蛋白含量的显著提高可能与芽孢杆菌、乳酸菌在生长中所分泌的各种蛋白酶有关[23]。田久东等[14]曾报道发酵西蓝花渣中的酸溶蛋白可较未发酵组提高150%以上,最高达到6.53%。而本研究中酸溶蛋白含量在发酵后西蓝花茎叶中可达8.41%,这可能与发酵菌种和发酵周期不同有关,需要进一步验证。最后,本研究中γ-氨基丁酸含量在代谢组和实际检测后均有显著提高,表明γ-氨基丁酸可作为一种潜在的标志物来分析发酵西蓝花茎叶品质,而更多差异小分子代谢产物的定量验证及其在动物生产中的作用还有待后续进一步研究探讨。
4 结论
本研究结果表明,西蓝花茎叶发酵前后共涉及35种差异代谢物且主要集中在蛋白质降解、氨基酸合成、植物次生代谢和维生素代谢等方面;发酵西蓝花茎叶中总酸、酸溶蛋白和γ-氨基丁酸的含量均显著增加,可能会提高其饲用品质。
