铁皮石斛(Dendrobium catenatum)叶艺形成机制的初探
2020-09-09郑宝强乔红娟李柏君KAOTachung
郑宝强,乔红娟,李柏君,KAOTachung,张 燕,王 雁*
(1. 国家林业局林木培育重点实验室,中国林业科学研究院林业研究所,北京 100091;2. Gao's Orchid Garden,AV Geroldo Azzoni 905 Rio Acima Jundiai SP,Brasil 13215-840;3. 北京市电气工程学校,北京 100025)
许多植物中都存在绿色叶片出现黄色或白色斑块的突变体[1-7],这些突变植株是研究光合作用、叶绿素合成代谢和叶绿体发育等过程的极佳材料[8]。此外,这类表型也是许多观赏花卉的主要观赏性状,是园艺工作者对花卉品种的主要改良和育种目标之一[9-11]。因此,探究叶片黄色或白色的斑化形成机制具有重要的生物学意义和应用价值。
叶绿素是使植物叶片显绿色的主要色素,其在植物体内的生物合成通路已经得到了详细描述和报道[12-15]。许多研究表明,叶绿素生物合成途径中结构基因的表达量变化会直接决定叶绿素在组织中的积累量,从而影响叶片颜色的表型[16-17]。当这些结构基因表达量下降时,会导致中间产物无法被催化成叶绿素,最终使叶片失绿从而产生白色或黄色的斑块或条纹[15,18-20];同时,造成组织失绿的具体机制在不同植物之间存在差异。因此,探索不同植物突变体叶片失绿的具体分子机制具有重要的意义,是对相关植物进行叶色园艺性状改良的理论基础和依据。
兰科(Orchidaceae)植物因其独特的花形、丰富的花色和迷人的花香而世界闻名,其许多种和栽培品种已成为广受大众欢迎的观赏花卉。叶片失绿的突变现象在兰花中被称为“叶艺”,其能够提高兰花的观赏价值,是兰花园艺工作者主要的选育目标之一[21]。但是,目前叶艺在兰科中的分子机制研究较少,仅在兰属(Cymbidium)[20,22-23]和卡特兰属(Cattleya)[24]有过相关报道,大多研究主要在生理水平上对叶艺机制进行探索[10,25-29]。近年来,许多石斛属(Dendrobium)的叶艺突变体在市场上越来越受欢迎。铁皮石斛(Dendrobium catenatum)在自然环境和人工栽培条件下常出现叶艺现象[30],这些叶艺铁皮石斛是石斛品种改良与培育的优质种质资源。此外,铁皮石斛的高质量基因组已经公布[31],这为研究铁皮石斛叶艺现象的分子机制提供了便利与基础。
1 材料与方法
1.1 实验材料
供试材料为铁皮石斛叶艺突变体的无性系,其叶片上有大面积白色斑块或条纹。本研究所用材料都培养在中国林业科学研究院温室中,采光为自然光照,正常栽培管理。
1.2 显微结构观察
1.2.1 叶片显微结构观察 分别取正常铁皮石斛(WT)、叶艺铁皮石斛叶子绿色组织(YYG)、叶艺铁皮石斛叶艺白色组织(YYW),用双面刀片快速切成薄片,放入有蒸馏水的玻璃皿中。选择其中最薄的透明度最大的薄片做成临时切片,在光学显微镜(Olympus,日本)下观察和拍照。
1.2.2 叶绿体超显微结构观察 根据Li等[15]的方法,将实验材料制备成切片;利用透射电镜(JEOL,日本)对切片中的叶绿体显微结构进行观察和拍照。
1.3 叶绿素及其前体物质含量的测定
本研究采用邵勤等[32]的叶绿素提取和含量计算方法,对不同处理称取等量实验材料并研磨至粉末,然后利用80%丙酮进行叶绿素萃取。通过UV2310Ⅱ分光光度计分别测定提取液波长在663、646、470 nm下的OD值。最后,根据相关计算公式得到不同实验处理的叶绿素含量。根据Bogorad[33]的方法对实验样品中胆色素原(Porphobilinogen,PBG)、尿 卟 啉 原 III (Uroporphyrinogen Ⅲ,UrogenIII)和粪卟啉 III(Coproporphyrinogen Ⅲ,CoprogenⅢ)进行提取和含量测定。根据Lee等[34]的方法对实验样品的原卟啉IX (Protoporphyrin IX,Proto IX)、镁原卟啉IX(Mg-Protoporphyrin IX, Mg-Proto IX)和原叶绿素酸酯(Protochlorophyllide,Pchlide)进行提取和含量测定。每处理进行3个生物学重复测定,并利用Omiscshare tools (http://omicshare.com/tools/)对数据进行Student's pairedt-test。
1.4 叶绿素合成相关基因的qRT-PCR定量
1.4.1 总RNA提取和质量检测 利用改良的SDS法[35]提取实验样品中的总RNA。利用NanoDrop 2000微量分光光度计(Thermo Scientific Inc., USA)对RNA的质量进行检测,OD260/280≥1.8、OD260/230≥1.8和浓度大于100 ng·μL-1的RNA被用于下一步的反转录。
1.4.2 相关基因的qRT-PCR定量 利用EasyScript One Step gDNA Removal and cDNA Synthesis SuperMix qPCR(全式金,中国)反转录试剂合成RNA的cDNA第一链。从已发表的铁皮石斛基因组[31]中找与叶绿素合成相关的基因,利用NCBI的Primer-BLAST进行引物设计(表1)。以Dc18S作为内参基因对数据进行标准化。根据TaKaRa TB GreenTMPremixEx TaqTM试剂盒(TaKaRa, Japan)使用说明书设置LightCycler480 System(Roche,USA)的qRT-PCR运行程序进行基因的qRT-PCR定量。每个基因在每个处理中进行3个生物学重复测定,并利用Omiscshare tools (http://omicshare.com/tools/)对数据进行Student's pairedt-test。

表1 叶绿素合成相关基因的qRT-PCR引物序列Table 1 The primer sequences of chlorophyll biosynthesis related genes for qRT-PCR
1.5 数据分析
利用SPSS 13.0与Excel 2010对生理指标进行统计和方差分析,基因定量采用Comparative CT(-ΔΔCt)法进行计算[36]。
2 结果与分析
2.1 叶艺突变体的显微观察比较
正常的铁皮石斛叶片为全绿色(图1a),而叶艺铁皮石斛的叶片上则出现了大面积的白色条纹或斑块(图1b、c)。对叶片横切面的显微观察结果显示:正常铁皮石斛(WT)和叶艺铁皮石斛叶片绿色组织(YYG)的部位中充满了绿色细胞(图1d、e),而叶艺铁皮石斛叶艺白色组织(YYW)的部位中则基本上看不到绿色细胞(图1f)。对这些组织细胞的超显微结构观察显示:WT和YYG的叶绿体呈饱满的半球形,其体内富含排列整齐的基粒片层和类囊体基粒,淀粉颗粒少(图2);而YYW的叶绿体则呈干瘪的三角形,体内基粒片层和类囊体基粒数量稀疏且排列无规则,淀粉颗粒多(图2)。这些结果显示叶子呈绿色的WT和YYG富含大量的绿色叶绿体且具有相似的叶绿体细胞结构,而YYW则与前两者相反。

图1 叶艺和正常铁皮石斛的形态特征和叶片显微观察Fig. 1 The morphological characteristics and microscopic structure of normal and mutant D. catenatum leaves
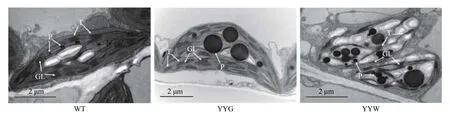
图2 石斛叶片叶绿体的超显微结构观察Fig. 2 The ultrastructure of chloroplast in D. catenatum leaves
2.2 叶绿素、叶绿素合成前体物质和类胡萝卜素的含量测定
叶绿素和类胡萝卜素含量测定显示:WT和YYG之间的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量无明显差异,但它们都显著高于YYW(图3)。这一结果显示YYW的叶绿素合成受到了阻碍,并支持了其白化表型特征和显微观察YYW中几乎没有绿色细胞的结果。
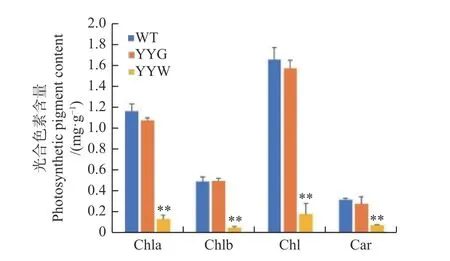
图3 野生型和突变型铁皮石斛叶片中光合色素的含量Fig. 3 Photosynthetic pigments content of WT and mutant leaves
叶绿素合成的上游中间代谢物中,YYW的胆色素原含量是WT的84%,而尿卟啉原Ⅲ和粪卟啉原Ⅲ含量则分别是WT的109%和137%(图4)。在叶绿素合成下游中间前体物质中,WT和YYG的原卟啉IX、镁原卟啉IX和原叶绿素酸酯含量是YYW的4.9~17.1倍,而它们二者之间的差距相较于YYW则较小(图4)。粪卟啉原Ⅲ是原卟啉IX和镁原卟啉IX重要合成前体[15]。因此,这些结果显示:YYW在合成叶绿素的过程中,原卟啉IX的合成受到了阻碍,中间产物无法正常流向叶绿素合成。
2.3 叶绿素生物合成途径中结构基因的表达量测定
WT、YYG和YYW中叶绿素生物合成途径中8个结构基因的qRT-PCR结果(图5)显示:YYW中的尿卟啉原脱羧酶(Uroporphyrinogen decarboxylase, DcHEME)、粪卟啉原氧化脱羧酶(Coproporphyrinogen III oxidase, DcHEMF)、原卟啉原氧化酶(Protoporphyrinogen oxidase, DcHEMG)和镁螯合酶H和镁螯合酶H亚基(Mg chelatase H subunit, DcCHLH)的表达量都显著低于WT和YYG。羟甲基后胆色素原合酶(Porphobilinogen deaminase, DcHEMC)的表达量在三者之间并无明显差异,而YYG和YYW中的胆色素原合酶(Porphobilinogen Synthase, DcHEMB)、镁螯合酶D亚基(Mg chelatase D subunit, DcCHLD)和镁原卟啉IX甲基转移酶(Mg protoporphyrin IX methyltransferase, DcCHLM)的表达量则都显著低于WT。值得注意的是,HEMF和HEMG是合成原卟啉IX的关键酶,而CHLH和CHLD则是合成镁原卟啉IX的关键酶[15]。
3 讨论
大多数植物的叶片为绿色,但许多植物都有叶片白化的突变体,而这种叶片出现白化的突变现象是许多观赏植物的重要观赏性状,具有很高的观赏价值[15]。叶绿素是植物进行光合作用的主要物质,同时它也是使植物组织呈现绿色的重要色素[37]。在许多植物的白化突变体中,其叶子的白化区域都出现了叶绿素积累量显著下降[13,38-40]。本研究通过显微观察发现,YYW基本没有绿色细胞,同时其叶绿体细胞结构畸形且体内基粒片层和类囊体基粒数量稀疏且排列无规则。许多研究表明,植物叶色的突变都伴随着叶绿体细胞结构的改变和体内类囊体基粒的无序排列[11,41-42]。因此,叶绿素含量的下降是导致叶艺铁皮石斛叶片出现白色条纹或斑块的主要原因。此外,粪卟啉原Ⅲ在YYW中的含量显著高于WT和YYG,而粪卟啉原Ⅲ的下游产物原卟啉IX、镁原卟啉IX和原叶绿素酸酯在WT和YYG中的含量是YYW的4.7~17.1倍。这进一步表明,YYW的叶绿素合成从粪卟啉原Ⅲ下游产物合成开始就受到了阻碍,而这可能是导致YYW中叶绿素无法正常合成的主要原因。
叶绿素生物合成途径中关键酶编码基因表达量的变化会直接影响叶绿素在植物组织的积累量,从而改变植物组织的颜色表型[15-17,43]。本研究中,叶绿素合成途径中的DcHEMF、DcHEMG、DcCHLH和DcCHLM在YYW中的表达量显著低于正常叶片。在叶绿素合成途径中,粪卟啉原Ⅲ经过HEMF和HEMG的催化后会生成原卟啉,然后分别通过CHLD、CHLH和CHLI进一步形成镁原卟啉IX[17]。值得注意的是,叶艺铁皮石斛中粪卟啉原Ⅲ含量显著高于正常叶片,而原卟啉和镁原卟啉IX含量则显著低于正常叶片。前人研究表明,金鱼草(Antirrhinum majus)叶片的黄绿表型是由Tam3插入使ChlH基因失去活性所导致[44]。水稻(Oryza sativa)叶片的突变体中,由于T-DNA插入到line 9-07117的镁螯合酶大亚基ChlH中,使其无法正常转录翻译,导致水稻叶片失绿[18]。银杏(Ginkgo biloba)叶片的黄化突变体中,GbPPO、GbCHLD、GbCHLH和GbCHLI在黄化叶片中的表达量下调,同时其粪卟啉原Ⅲ含量显著高于正常叶片,而原卟啉IX和镁原卟啉IX的含量则相反[15]。本研究中,DcHEMF、DcHEMG和DcCHLH在YYW中的低表达使粪卟啉原Ⅲ无法正常流向叶绿素合成,造成叶绿素的积累量减少,最终导致叶片失绿。这些研究结果为今后石斛属的叶艺性状改良提供了理论依据和分子基础。
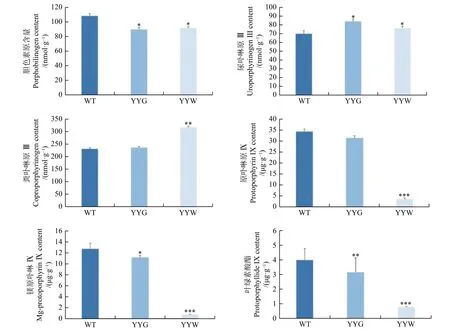
图4 野生型和突变型铁皮石斛叶片中叶绿素合成的中间产物含量Fig. 4 The content of chlorophyll intermediates in WT and mutant leaves
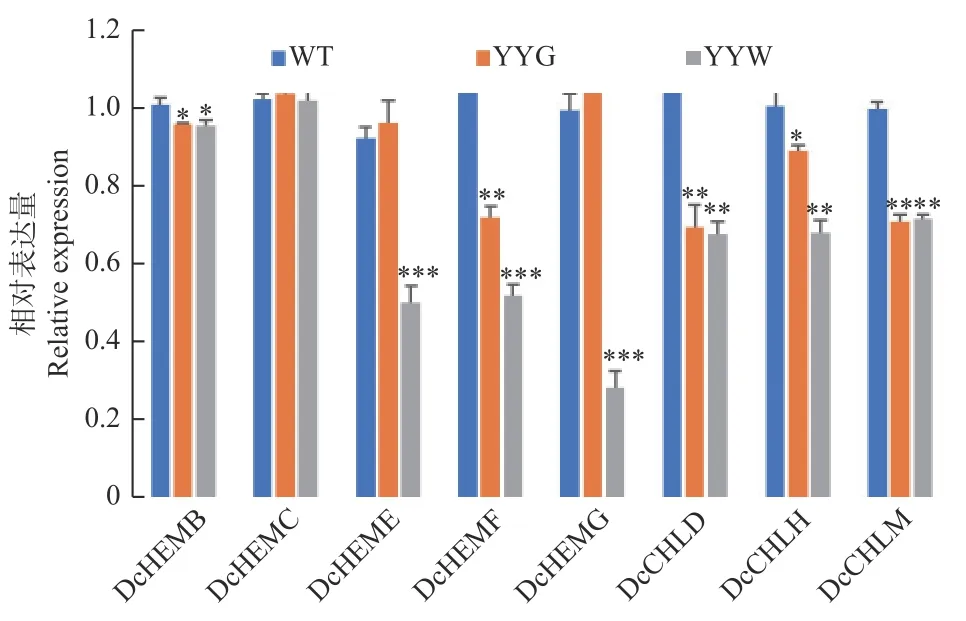
图5 野生型和突变体石斛叶片中叶绿素合成相关基因的表达量变化Fig. 5 The relative expression of some chlorophyll biosynthesis relative genes in WT and mutant leaves
4 结论
DcHEMF、DcHEMG和DcCHLH在叶艺铁皮石斛叶片白色区域中的低表达使粪卟啉原Ⅲ无法正常流向叶绿素合成,这导致了叶绿素含量的下降,是使铁皮石斛出现叶艺表型的主要原因。
