双线紫蛤COI基因序列多态性与水环境分析
2020-08-21王朋云
王朋云
(1.莆田市水产技术推广站,福建 莆田351100;2.福建省海洋生物增养殖与高值化利用重点实验室,福建 厦门361013)
双线紫蛤(Soletellinadiphos)常埋栖于潮间带中、下区至浅海的细沙和砂泥质海底,分布地区为印度洋及西太平洋,在中国主要产于福建以南沿海。双线紫蛤肉质鲜美,食用价值和经济价值高,但天然采捕产量较少,已被列入福建省重点保护野生动物名录。南日岛是福建沿海双线紫蛤栖息与繁殖的自然主产区,亦是南日群岛的主岛,位于平海湾与兴化湾的交汇处,呈西北-东南方向具芒角哑铃状排列,长约14.0 km,最狭处宽约3.0 km,岛岸线总长66.43 km,面积45.08 km2,该海域分布的双线紫蛤资源常被供应于青岛、福州、厦门多家科研院所高校做研究和平潭、连江等地水产企业做苗种繁育。海区生物资源的丰度与遗传多样性、海域生态环境的状况存有紧密的联系,为遏制野生双线紫蛤种群资源衰退,聚焦当地种质遗传背景并清晰了解生境水域条件,开展人工繁殖育苗,评估生态影响科学增殖放流以复壮种群,进行群体内基因序列多态性及海区水样检测结果统计分析势在必行,对保护和修复南日岛的双线紫蛤种质资源意义重大。目前国内已有双线紫蛤的研究报道,主要在人工养殖扩繁[1-3]、形态差异[4]、软体部营养成分[5]、血细胞组成[6]和系统亲缘进化[7-8]等方面,但尚未有开展双线紫蛤地理群体序列多态性和栖息地水环境分析的详细报道。
线粒体COI基因序列被广泛应用于检测双壳贝类等的群体遗传多样性、遗传结构和系统发育研究,基因序列的单核苷酸多态性(Single nucleotide polymorphism,SNP)位点是可被应用于区别不同地理种群以及评估自然群体遗传变异程度的有效分子标记。开展海域水体盐度、温度、pH、溶解氧、叶绿素a、透明度常规和化学需氧量、无机氮、无机磷、石油类常见污染物等多项参数追踪检测,是全面掌握海区水环境状况动态变化特征的科学技术手段。为调研双线紫蛤南日岛自然种群的遗传差异和生长环境情况,本研究在直接利用COI条形码双向测序检测SNP位点的基础上,结合统计多年南日岛海域水样检测数据,对其遗传多样性与生长海域水环境情况进行分析,以期为双线紫蛤的遗传标记、生态调查等研究积累数据。
1 材料与方法
1.1 材料
实验所用双线紫蛤野生群体样本由福建省南日岛海洋生物技术有限公司在南日岛海域所采集,同一海区水样检测结果由莆田市海洋与渔业环境监测站提供。
1.2 方法
1.2.1 序列多态性
于2018年10月22日—11月28日捕获的双线紫蛤活体立即低温带回实验室处理,随机抽选30枚个体,剪取前闭壳肌抽提基因组DNA,检测定量后低温保存。引物(表1)为依据简并引物[9]扩增测序结果在通用引物[10]基础上修改下游碱基而来,由生工生物工程(上海)股份有限公司合成,PCR扩增COI序列体系及条件同《双线紫蛤(Soletellinadiphos)形态、序列测定和同工酶的初步研究》采用的方法[7],回收产物后送样。样本双向直接测序结果经人工校对后采用NCBI blastn在线比对验证核酸序列同源性、BankIt程序批量提交至GenBank,BioEdit编辑,Clustal X 对位,MEGA 6.0转换格式、统计碱基组成和转换颠换比、Kimura 2-parameter计算遗传距离(bootstrap值重复1 000次),DnaSP 6[11]计算多态位点数目、单倍型数目、单倍型多样性(Hd)、核苷酸多样性(Pi)、平均核苷酸变异数(k)及Tajima’s D[12]和Fu’s Fs中性检验值[13]。

表1 双线紫蛤COI基因序列片段扩增所用的引物
1.2.2 栖息地水环境
2008—2018 年选择数个具有季节代表性的月份(表2)安排出海取样,南日岛的双线紫蛤栖息海区初期(2008—2010年)采样站点6个,后续(2011—2018年)追加调整为12个,所布设的12个站点位置同2019年8月10—16日监测站位[14](图1),10项理化因子样品的采样、运输、测定与质量控制过程均严格依据GB 17378—2007《海洋监测规范》要求进行[15]。水样的盐度、温度、pH、溶解氧、叶绿素a、透明度、化学需氧量、无机氮、无机磷、石油类检测结果按航次整理分项记入Excel软件,计算不同时间尺度下的极值、均值、标准偏差并绘制月份数据点折线图。

表2 南日岛海区水样采集时间
2 结果
2.1 序列多态性
双线紫蛤样本编号为1~50(图2-A),盲选出30枚个体(图2-B)进行PCR,扩增产物电泳结果与预期目的序列长度一致(图2-C),测得的30条COI基因序列片段(MN176021~MN176050)分子量为702 bp,去除上、下游引物后为655 bp,其中保守位点642个,群体序列的4种碱基组成:T>G>A>C,个体间的各碱基含量变化不大,C占比最低、为16.07%,T比例最高、为41.95%,平均(G+C)含量39.52%明显低于(A+T)含量60.48%(表3),样本间的遗传距离介于0~0.009 2,总平均距离为(0.002 9±0.001 0)。

表3 双线紫蛤COI基因序列片段碱基组成
双线紫蛤群体30枚个体的COI片段中共计检测出13个变异位点(表4),出现频率为1.98%,序列间的变异位点分布不均匀,在68~201 bp、393~532 bp两段相对保守的区域没有突变。其中202位点、601位点的突变频率大于25.00%,分别为30.00%、26.67%,202位点突变模式为T/C转换(图3-A),601位点为G/A转换(图3-B),这2个SNP位点的波峰均较整齐且无杂峰干扰。全部变异位点经双向测试为纯合突变,简约信息位点4个(202、203、265、601),自裔位点9个,无插入/缺失位点。在这13个位点中仅有1个颠换位点(占7.69%),即55位点的A/T颠换,其余为转换位点,T/C之间转换5个、C/T转换3个、G/A转换3个、A/G转换1个,T/C转换比例较高、占41.67%;序列中出现的突变总数为34,转换/颠换的偏倚率(R)为13.04(>2.0)。群体的序列翻译成氨基酸,发现203位点的C/T转换引起了相应氨基酸亮氨酸(Leu)/苯丙氨酸(Phe)的变化,为非同义替换,其余12个位点属于同义替换,非同义和同义替换比值为0.08(<1.0)。编码氨基酸的密码子中,第3位的转换最多、有9个位点,第1位次之、有3个(203、392、533),颠换出现在第3位、有1个位点,第2位的转换和颠换数为0。

表4 双线紫蛤COI基因序列片段检出的SNP位点
双线紫蛤群体30枚个体的COI序列依据片段中SNP位点的分布情况进行统计,共检测到9个单倍型(表5),占样本总数的30.00%,单倍型多态性指数(Hd)为(0.598 0±0.103 0)(Hd>0.5),核苷酸多样性指数(Pi)为(0.002 9±0.000 6)(Pi<0.005),平均核苷酸变异数(k)为1.912 6,中性检验Tajima’s D值为-1.377 5(<0.00)、不显著(P>0.05),Fu’s Fs值-2.354 0(<0.00)、不显著(P>0.05)。单倍型Hap1包含的样本个体数最多、为19枚(样本1、2、4、7、10、14、19、22、25、27、28、29、31、32、35、39、43、45、47),占样本含量的63.33%,是群体基于COI序列片段的主要单倍型,Hap5次之、含3枚(样本12、41、42),Hap4有2枚(样本11、15),其余Hap2(样本5)、Hap3(样本6)、Hap6(样本13)、Hap7(样本24)、Hap8(样本36)、Hap9(样本46)6个单倍型各仅出现1枚个体,占单倍型种类的66.67%。
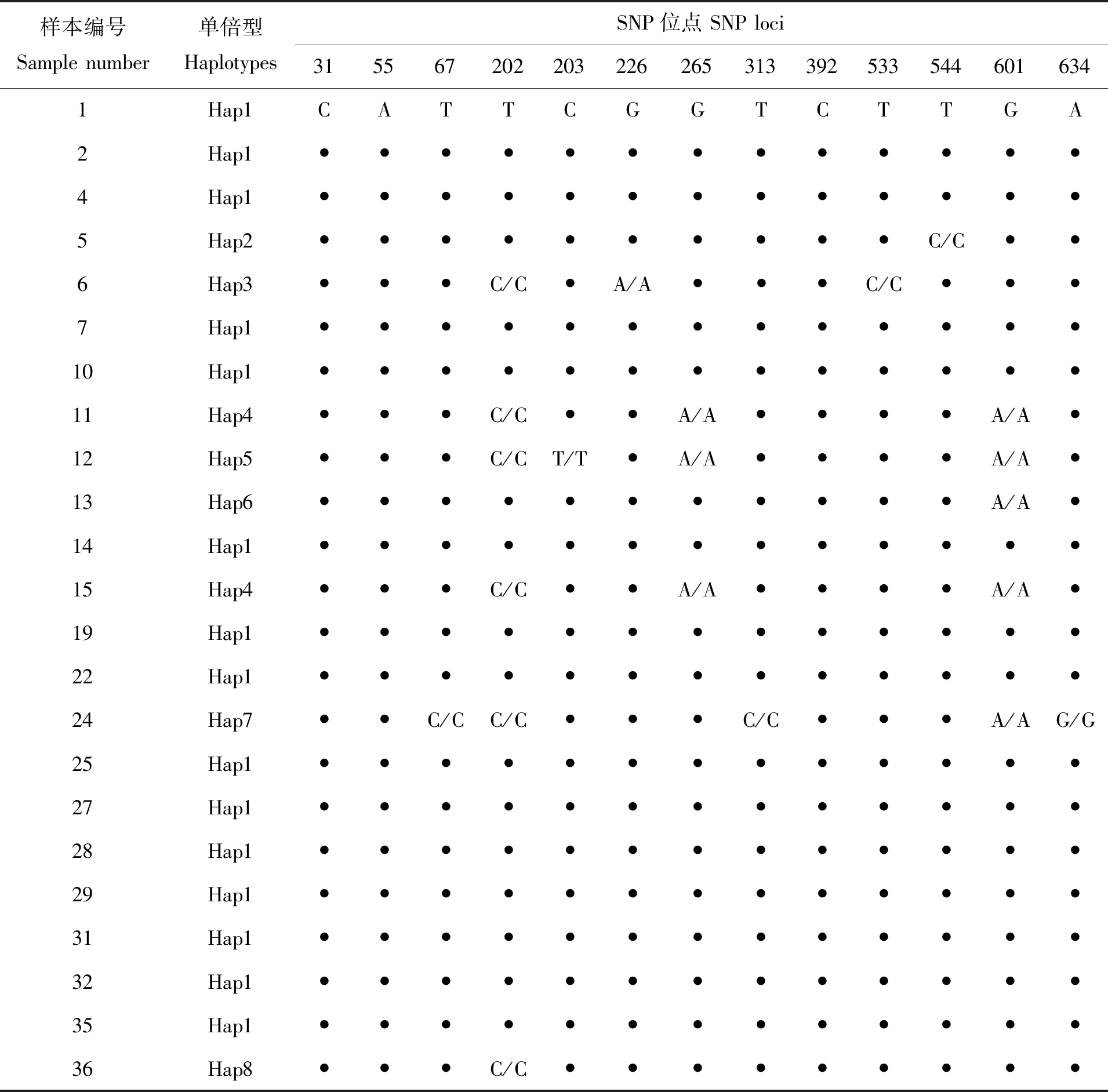
表5 双线紫蛤COI基因序列片段单倍型和SNP位点的分布情况

续表5
2.2 栖息地水环境
2008年4月—2018年12月南日岛双线紫蛤栖息海域74个航次海水的10项理化因子共测得7 740个水环境数据,其中盐度、温度、pH、溶解氧、叶绿素a、透明度、化学需氧量、无机氮、无机磷和石油类分析项目检测结果各774个。
2008—2018年海水盐度变化范围为25.80~34.90,11年间的月平均波动为26.84~34.33,平均值为(31.48±1.89),盐度总体上夏季(33.60±0.40)>春季(31.40±1.17)>秋季(30.07±1.54)>冬季(29.11±1.22),在秋季变化幅度较大;各年盐度春、夏、秋三季变化均呈现出先上升后下降的趋势,夏季均为全年的最高值,春季除2013年外均为次高值,有8年最低值出现在冬季、7年次低值出现在秋季,年平均值:2018年>2008年>2011年>2010年>2015年>2013年>2017年>2009年>2014年>2012年>2016年;最低值出现在2016年10月的站点当中,当月平均为(27.46±0.59),最高在2016年7月,当月平均值为(34.33±0.29)。
2008—2018年海水温度变化范围为12.80~29.90℃,11年间的月均波动为13.27~29.33℃,平均值为(21.73±4.83)℃,温度总体上夏季(27.71±1.09)℃>秋季(23.33±1.54)℃>春季(18.72±3.26)℃>冬季(16.89±2.19)℃,在春季变化幅度较大;除2010年缺少冬季数据外,各年温度四季变化均呈现出先上升后下降再下降的趋势,温度全年的最高值均出现在夏季,次高值均出现在秋季,有8年最低值出现在冬季、9年次低值出现在春季,年平均值:2008年>2015年>2017年>2010年>2018年>2016年>2014年>2013年>2011年>2012年>2009年;最低值出现在2012年3月,当月平均为(13.27±0.43)℃,最高在2012年8月,当月平均值为(29.33±0.33)℃。
2008—2018年pH变化范围为7.73~8.74,11年间的月均波动为7.89~8.54,平均值为(8.09±0.09),pH总体上春季(8.13±0.09)>冬季(8.10±0.06)>秋季(8.09±0.06)>夏季(8.03±0.09),在春、夏两季变化幅度相对较大;各年pH春、夏两季变化均呈现出下降的趋势,有6年最高值出现在春季,8年夏、秋两季升高且最低值出现在夏季,6年秋、冬两季升高或次低值出现在秋季,年平均值:2010年>2011年>2008年>2018年>2012年>2017年>2009年>2014年>2013年>2015年>2016年;最低值出现在2016年7月,当月平均为(7.92±0.08),最高在2010年5月,当月平均值为(8.54±0.13)。
2008—2018年溶解氧变化范围为5.45 ~9.92 mg/L,11年间的月均波动为5.64 ~9.31 mg/L,平均值为(7.44±0.71)mg/L,溶解氧总体上春季(7.89±0.61)mg/L >冬季(7.49±0.66)mg/L >秋季(7.30±0.40)mg/L >夏季(6.84±0.54)mg/L,在冬季变化幅度较大;各年溶解氧含量春夏变化均呈现降低的趋势,有8年最高值出现在春季,9年夏、秋两季升高且最低值出现在夏季,6年秋、冬两季升高,7年次低值出现在秋季,年平均值:2012年>2014年>2017年>2010年>2018年>2013年>2016年>2011年>2015年>2009年>2008年;最低值出现在2009年7月,当月平均为(5.64±0.19)mg/L,最高在2010年5月,当月平均值为(9.31±0.67)mg/L。
2008—2018年叶绿素a变化范围为0.06 ~20.93 μg/L,11年间的月均波动为0.33 ~15.57 μg/L,平均值为(1.81±1.84)μg/L,叶绿素a总体上夏季(2.57±1.79)μg/L >春季(1.75±2.18)μg/L >秋季(1.54±0.99)μg/L >冬季(0.76±0.26)μg/L,在春季变化幅度较大;除2010年外,其余各年叶绿素a含量春、夏两季变化均呈现出升高,秋、冬两季则均降低的趋势,有9年夏、秋两季降低,8年最高值出现在夏季,9年最低值出现在冬季,年平均值:2010年>2009年>2008年>2011年>2017年>2016年>2012年>2013年>2015年>2018年>2014年;最低值出现在2018年3月,当月平均为(0.33±0.27)μg/L,最高在2010年5月,当月平均值为(15.57±4.38)μg/L。
2008—2018年透明度变化范围为0.30 ~2.00 m,11年间的月均波动为0.40 ~1.79 m,平均值为(0.79±0.24)m,透明度总体上春季(0.86±0.24)m >夏季(0.83±0.26)m >秋季(0.73±0.16)m >冬季(0.61±0.17)m,在夏季变化幅度较大;各年透明度有8年春、夏两季降低且最高值出现在春季,7年夏、秋两季或秋、冬两季降低,年平均值:2008年>2014年>2010年>2011年>2013年>2009年>2015年>2016年>2017年>2018年>2012年;最低值分别出现在2009年3月、2012年12月、2017年12月的站点,当月平均依次为(0.45±0.08)m、(0.45±0.16)m、(0.40±0.04)m,最高在2008年7月,当月平均值为(1.79±0.13)m。
海水年盐度、年叶绿素a含量大多数月份的数值随着水温升降,月均波动趋势同温度变化相对较一致;溶解氧含量则一般随着温度升高而降低,检测结果其变动与水温升降呈现出一定的负相关;pH、透明度的季节月份变化没有规律,二者波动与温度变化的直接影响关系不明显(图4)。
2008—2018年南日岛海区的化学需氧量变化范围为0.11~2.28 mg/L,11年间的月平均波动为0.22 ~1.59 mg/L,均值为(0.67±0.29)mg/L,化学需氧量总体上冬季(0.72±0.36)mg/L >夏季(0.68±0.28)mg/L >春季(0.67±0.28)mg/L >秋季(0.58±0.21)mg/L,在冬季变化幅度较大;各年化学需氧量季节变化无规律,年均值:2010年>2018年>2009年>2008年>2017年>2013年>2014年>2011年>2016年>2015年>2012年;最低值出现在2008年9月的站位,当月平均值为(0.22±0.07)mg/L,最高在2010年5月,当月平均值为(1.22±0.55)mg/L。
2008—2018年无机氮变化范围为0.03~0.54 mg/L,11年间的月均波动为0.04~0.45 mg/L,均值为(0.23±0.12)mg/L,无机氮总体上冬季(0.37±0.09)mg/L >秋季(0.30±0.08)mg/L >春季(0.23±0.10)mg/L >夏季(0.13±0.07)mg/L,在春季变化幅度较大;各年无机氮含量春、夏、秋三季变化均呈现出先减少后增加的趋势,最低值均在夏季,年均值:2016年>2012年>2014年>2013年>2017年>2015年>2009年>2008年>2011年>2018年>2010年;最低值出现在2017年7月,当月平均值为(0.04±0.01)mg/L,最高在2016年4月,当月平均值为(0.36±0.09)mg/L。
2008—2018年无机磷变化范围为0~0.37 mg/L,11年间的月均波动为0.00~0.16 mg/L,均值为(0.02±0.03)mg/L,无机磷总体上冬季(0.03±0.01)mg/L >秋季(0.03±0.02)mg/L >春季(0.02±0.02)mg/L >夏季(0.01±0.03)mg/L,在夏季变化幅度较大;除2008年外,各年无机磷含量夏、秋两季变化均呈现出增加的趋势,有9年春、夏两季减少的波动,最低值出现在夏季,年均值:2008年>2010年>2013年>2012年>2016年>2011年>2018年>2009年>2014年>2017年>2015年;最低值出现在2017年7月,当月平均值为(0.01±0.00)mg/L,最高在2008年6月,当月平均值为(0.16±0.14)mg/L。
海水年化学需氧量的测试结果:季节月份变化没有显著特点,不与海区水温的升降同步或相反;无机氮的部分年度数据表现出了随着水温升高而降低、降低而回升的负相关趋势,部分月份的结果则不支持这一现象;无机磷含量的月均数值波动同温度变化的关系存在着和无机氮检测结果类似的负相关情况(图5)。
2008—2018年南日岛海区的石油类检测数据变化范围为0~0.49 mg/L,11年期间的月平均波动为0~0.20 mg/L,平均值为(0.02±0.03)mg/L,石油类总体上夏季>秋季>春季>冬季,在夏季变化幅度较大;各年石油类含量季节变化无规律,年平均值:2009年>2008年>2016年>2014年>2013年>2018年>2017年>2011年>2015年>2012年>2010年;最低值在2008年12月、2011年12月、2013年5月的站位中都有出现,当月平均值皆为0 mg/L,最高在2009年7月,当月平均值为(0.20±0.22)mg/L。石油类含量多年度的月份波动不规律,与海水的温度高低变化模式没有发现明显的关联(图6)。
11年间南日岛双线紫蛤群体栖息海区水环境的盐度、温度等10项理化参数检测结果年际统计分析均没有出现逐年递增或递减的波动现象(表6)。
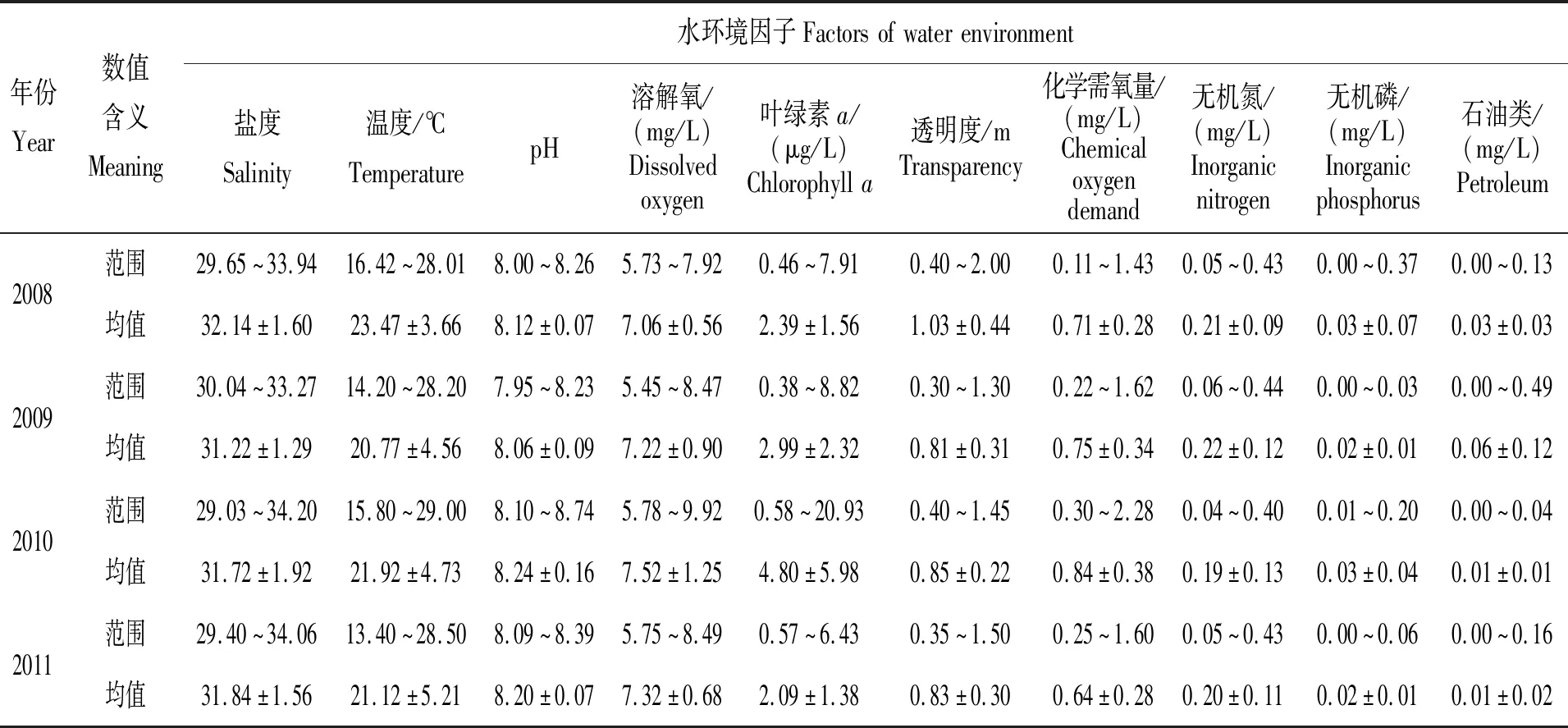
表6 南日岛海区水环境各项理化因子历年比较

续表6
3 讨论
3.1 序列多态性
本研究利用PCR产物直接测序法对双线紫蛤群体的COI基因片段进行分析,在样本个体的标记序列PCR过程中,简并引物[9]能够很好地扩增,但因其正、反向引物皆较多简并碱基而不适用于直接测序,需经连接转化;而通用引物[10]一直无法成功扩增测序,表明其通用性不适用于双线紫蛤。通过简并引物的单克隆测序结果,发现产物重叠区域中不含通用引物的上游引物(LCO1490)结合位点,而下游引物(HCO2198)锚定区域26个碱基中分布着6个变异位点,据此修改后的引物则可以有效地应用于双线紫蛤群体样本的COI基因部分序列测定。群体序列间的碱基组成含量,(A+T)含量占比优势明显,与菲律宾蛤仔(Venerupisphilippinarum)[16]、文蛤(Meretrixmeretrix)[17]、丽文蛤(M.lusoria)[18]、西施舌(Coelomactraantiquata)[19]等海水双壳类COI基因序列的碱基组成特点相似。双线紫蛤样本个体间的遗传距离都很小,遗传距离的大小往往与分化的程度正相关,说明南日岛群体的遗传分化程度不高。转换/颠换比值远大于临界值2.0,COI序列的突变未达到饱和状态,可有效检测多样性,转换比颠换的发生频率明显偏高,表明双线紫蛤群体片段区域内的变异相对较多的是发生在同类型碱基之间,群内个体间的亲缘关系较近,符合近缘阶元间核苷酸替换以转换为主的常规[20-21]。获得的测序峰图碱基主峰清晰,无干扰杂峰,易于对COI基因片段的突变位点进行精确地统计分析,突变大多数发生在密码子第3位,占76.92%,第1位和第2位的突变较低,分别占23.08%和0%,392位点、533位点虽同203位点一样突变发生在密码子第1位,但CTA/TTA、TTG/CTG在无脊椎动物线粒体基因中都表达为亮氨酸(Leu),故碱基替换未引起氨基酸的变化。群体序列的非同义和同义替换比值小于1.0,说明COI基因在进化过程中可能受到负选择压力的作用。在30个样本的655 bp序列中检测出了13个突变位点,变异位点之间的位置分布、发生突变情况和突变频率暂未观察到明显的规律,只在小部分样本中具有一定程度的相似,但并不总是一致。遗传多样性的高低直接体现了群体的遗传变异程度[22],也间接反映了其适应环境变化能力的强弱[23-25],遗传变异低的群体面对生境变化的压力也较大[26],种群的遗传多样性常用单倍型及核苷酸多样性指数[27-28]来衡量,变异多样性程度与指数大小正相关。南日岛群体的Hd略大于标准值0.5、单倍型多样性相对稍微偏高,而Pi低于0.005、核苷酸多样性较低,低于菲律宾蛤仔(V.philippinarum)[16]、光滑河蓝蛤(Potamocorbulalaevis)[29]、奇异指纹蛤(Acilamirabilis)和指纹蛤(A.divaricata)[30]等海洋贝类,显示单倍型与核苷酸多样性高低没有直接关系,说明双线紫蛤遗传多样性水平较低,对生存水环境条件改变敏感,种群可能有面临严重衰退的风险,有必要加强保护,这与曾广泛分布于南日岛周边海区的双线紫蛤野生资源量近些年受干扰分布范围有所减小的情况相符,遗传多样性因群体变小而降低。种群的高Hd低Pi被认为可能是在经历过数量急剧减少的瓶颈效应之后,幸存的优势个体得以繁殖形成较小的种群,短时间内积累的核苷酸多样性较低,群内个体序列所属的单倍型之间的差异较小[31-32]。Tajima’s D检验值与Fu’s Fs检验值为负,但二者的统计检验P值并未达到显著性差异水平,即没有明显地偏离0,说明南日岛群体符合中性进化模型,显示在过去较近的历史时期内群体大小保持稳定状态,内部的遗传分化现象还不够明显。线粒体基因一般遵循母系遗传[33],根据单倍型数量,推测抽选测序的30枚个体至少是9枚雌性亲贝自然产卵繁殖的子代,单倍型Hap1代表的母系亲本集体在群体中拥有最高的出现频率,较其他8个单倍型,对本地海区生境有着更强的适应能力,生存优势明显。本研究因条件所限仅采集到莆田南日岛1个地理群体,数次联系赴广东[4]、长乐[6]等地搜集样本未能如愿获取,而鉴于已有资料指出双线紫蛤在我国主要分布于福建以南沿海,期待后续能够结合其他海区的自然群体进行遗传分化和遗传结构等方面的分析。
3.2 栖息地水环境
现有年际3—12月的水样检测数据表明,双线紫蛤的分布和生长发育对水环境有一定的要求,能够适应南日岛潮间带及浅海海区月均盐度26.84~34.33的波动幅度,适应盐度区间与中国紫蛤(Hiatulachinensis)[34]、波纹巴非蛤(Paphiaundulate)[35]对盐度的耐受范围有大部分重合,明显不同于主要生活在盐度相对较低的江河口咸淡水交汇处的尖紫蛤(H.acuta)[36]。栖息地环境水温13.27~29.33℃的变化范围,显示出双线紫蛤对水环境温度的适应能力上限高于紫石房蛤(Saxidomuspurpuratus)[37],低于皱纹蛤(Periglyptapuerpera)[38]但比其耐低温,与大西洋浪蛤(Spisulasolidissima)[39]的习性相当。每月取样检测日期因阴雨天气、潮汐或任务调整而间隔时间没有固定一致,个别月份如每年的1—2月受低温或海况不佳风急浪大影响无法出海,同一海区的水质浮标24 h全天候在线监测设备[40]绘制的趋势图显示2019年1—2月昼夜水温变幅区间在10.02~13.64℃,说明双线紫蛤实际上可以短时间内耐受生长环境10.00℃以上的低温。2008年8月水温低于邻近的7月、9月,也低于其他所有年份的同期值,海区表层水温的日变化受太阳辐射、气象条件等的调节[41],日间水温逐渐上升,一般在下午至傍晚的时刻达到最高值[42],夜间水温呈下降的趋势,凌晨至日出前的时刻达到最低值[43],反映出之后根据天气考虑合理安排出海时间对水温测量结果至关重要;10月水温在11年间的同期数据中处于第二高位,11月采样时间与上月较近,冷空气还未到达而未造成降温,温度略高0.26℃,但尚在正常逐日变化差值波动范围内。2010年5月海水的叶绿素a含量检测结果偏高,约是2017年7月第二高值6.34 μg/L的2.46倍,另外显微镜检在水样中发现大量的东海原甲藻(Prorocentrumdonghaiense),应该是该藻的快速生长繁殖导致了叶绿素a含量猛增。透明度的降低,除了水温、营养等条件适宜浮游生物活跃增殖的影响外,亦会受海区水深[44]、气象风浪[45]翻动海区底泥沉积物或悬浮物[46]等的影响。冬季低温条件下细菌、浮游植物等生长繁殖活力不强,初级生产力水平降低对营养元素吸收利用减弱,使得南日岛海域的无机氮、无机磷在冬季含量较高,多年月均变化情形基本一致,与福建多地沿海海水氮和磷监测信息的三季时空消长特征相似[47-48]。石油类含量数值在2009年7月出现异常,达到2008年7月次高值0.06 mg/L的3.33倍,各站点的最低检测值为0.03 mg/L,最高为0.49 mg/L,差异较大,可能与采样雇佣船只自身或个别站位附近恰逢有作业航道有一定的关系。通过统计南日岛海域水样的各项理化因子检测数据,分析相应参数的水平或浓度多年度季节月份均值波动变化范围,对双线紫蛤栖息的水环境状况进行初步调查了解,为进一步的生态习性观察实验提供了参考。
致谢:实验得到福建省水产研究所曾志南研究员及巫旗生助理研究员、莆田市海洋与渔业环境监测站张丽高级工程师及全体同仁、福建省南日岛海洋生物技术有限公司陈珍赐总经理的诸多支持,特此致谢。
