谷氨酰胺和丙酮酸钠对NK-92 细胞体外扩增的影响
2020-08-07应丹妮陈焕芸蔡海波谭文松
应丹妮, 陈焕芸, 付 岩, 蔡海波, 谭文松
(华东理工大学 生物反应器工程国家重点实验室, 上海 200237)
1 前 言
免疫效应细胞由于其独特的杀瘤能力和治疗优势使其在肿瘤疾病治疗中备受瞩目[1-2]。在治疗过程中,为了获得满意的临床效果,需要分离患者自体免疫效应细胞通过体外扩增技术获得符合临床要求的细胞数量。由于从肿瘤患者体内难以采集大量免疫效应细胞,且收获的细胞质量受患者身体状况影响很大,常常无法实现有效扩增,不仅使患者遭受巨大痛苦,也影响治疗效果[3-4]。因此,如何提高免疫细胞体外扩增效果并维持其杀伤活性是亟待解决的问题[5-7]。
氨基酸作为哺乳动物细胞培养基中的重要成分,其不仅是细胞增殖所需生物大分子的合成原料,某些氨基酸也可作为能源物质为细胞体外扩增提供ATP,除此之外,还可以作为嘌呤、嘧啶、氨基糖和抗氧化剂合成的底物,参与其他生物合成途径去维持细胞的完整和功能[8]。研究发现,当免疫效应细胞被激活时,抑制氨基酸转运蛋白的表达从而阻碍氨基酸的摄取时,会抑制它的扩增和功能[9-13]。同时丙酮酸是多种氨基酸合成的起始化合物,免疫效应细胞受到刺激时会通过丙酮酸羧化酶加速丙酮酸氧化以促进免疫细胞的增殖[14]。由此可见,优化培养基中氨基酸种类和浓度是免疫细胞体外扩增过程中的关键环节。而有血清培养基成分复杂,现有商业化无血清培养基中的氨基酸浓度通常也是保密的,无法使用现有的培养基进行优化以适满足细胞需求[15]。
为此,本文以具有CD3-CD56+表型特征和高细胞毒性的自然杀伤细胞系(natural killer-92 cells, NK-92)细胞为对象[16],在一款自主设计的化学成分简单、明确的无血清培养基SFM-MIN 中分析了NK-92 体外扩增前期氨基酸的消耗,探查关键营养组分,采用正交实验确定其最适浓度。并在有/无血清培养体系中,以细胞活性、总细胞扩增、培养物中CD3-CD56+细胞比例、胞内ATP 含量和细胞杀伤活性为指标,验证了研究结果。为后续开发免疫效应细胞高效扩增的无血清培养基提供了技术支持和依据。
2 材料与方法
2.1 实验材料
NK-92 细胞来源于人恶性非霍奇金淋巴瘤患者外周血,购自南京科佰生物科技有限公司;α-MEM 培养基购自美国Thermo 公司;HIPP-T009 淋巴细胞无血清培养基购自上海倍谙基生物科技有限公司,其谷氨酰胺和丙酮酸钠浓度分别为3.3 和1.0 mmol·L-1;胎牛血清购自美国Hyclone 公司;马血清购自以色列Biological Industries 公司;白细胞介素2(IL-2)购于美国Peprotech 公司;鼠抗人FITC-CD3 抗体和鼠抗人PE-CD56 抗体购自美国 Becton Dickinson 公司;二甲基亚砜购自美国 Merck 公司;丙酮酸钠购自美国Applichem 公司;谷氨酰胺购自美国Merck 公司;CellTiter-LumiTMPlus 发光法细胞活力检测试剂盒购自碧云天生物科技有限公司。SFM-MIN 是自配的无血清培养基,其谷氨酰胺和丙酮酸钠浓度分别为0.0 和0.0 mmol·L-1。
2.2 方法
2.2.1 细胞培养
NK-92 细胞以 5×105cells·mL-1的密度接种于添加 12.5% 胎牛血清和 12.5% 马血清的 α-MEM 培养基或无血清培养基T009 中,并添加1 000 U·mL-1IL-2,在37 ℃、5% CO2的饱和湿度培养箱中培养15 d。培养过程中,每天取样计数,并补充新鲜培养基和IL-2,调回最初接种密度。
2.2.2 细胞冻存
将处于对数生长期的NK-92 细胞以1 000 r·min-1离心5 min,弃上清后悬浮于由90% 胎牛血清和10%二甲基亚砜组成的冻存液中,以1×107cells·mL-1的密度接入1.5 mL 的冻存管,每管1 mL,4 ℃ 放置30 min,-20 ℃ 放置2 h 后转移到 -80 ℃ 冰箱中,24 h 后放入液氮中冻存7 d。
2.2.3 细胞表型分析
收集(5~10)×105个细胞,加入抗体稀释液1 mL,1 500 r·min-1离心5 min,洗涤2 次后,将细胞重悬于 100 μL 抗体稀释液中。分别将 8 μL 浓度为 0.2 mg·mL-1FITC-CD3 抗体和 8 μL 浓度为 0.2 mg·mL-1PE-CD56 抗体加入至上述细胞悬液中,4 ℃ 孵育30 min 使细胞表面抗原与加入的抗体充分结合,最后加入抗体稀释液1 mL,1 500 r·min-1离心5 min 除去未结合的抗体并将细胞重悬于300 μL 抗体保存液中,采用FACS Aria I 流式细胞仪(美国Becton Dickinson 公司)分析培养物中CD3-CD56+细胞的比例,结果用FlowJo 软件进行处理和分析。
2.2.4 胞内ATP 含量测定
采用化学发光方法测定胞内ATP 含量,取一定量的待测细胞,制备密度为1.5×104cells·mL-1的细胞悬液,将100 μL 细胞悬液加入到事先加有100 μL·孔CellTiter-LumiTMPlus 发光检测试剂的96 孔板中,以添加100 μL 培养基为对照,采用荧光酶标仪进行化学发光检测。
2.2.5 NK-92 细胞对K562 杀伤活性的检测
以NK-92 细胞为效应细胞(E),以处于对数生长期的K562 细胞为靶细胞(T),利用2 个细胞共培养时NK-92 细胞对K562 细胞的杀伤作用评价NK-92 细胞的杀伤活性。分别将1×105个NK-92 细胞(E)、1×104个K562 细胞(T)、以及上述2 种细胞的混合物(ET)接种于装有100 μL 无血清培养基的96 孔板中,置于37 ℃,5% CO2培养箱共培养4 h 后每孔加入10 μL CCK-8 试剂,37 ℃ 孵育2 h 后用酶标仪检测效应细胞(E)、K562 细胞(T)和 2 种细胞的混合物(ET)在 450 nm 处的吸光度 ODE、ODT和 ODET。按式(1)计算 NK-92细胞对K562 细胞的杀伤活性:
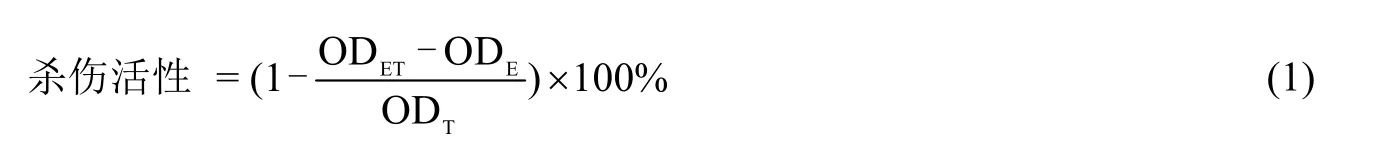
2.2.6 氨基酸浓度分析
采用浓度已知的20 种氨基酸混合液标准品稀释制成不同浓度的氨基酸标准品混合液,以邻苯二甲醛一氯甲酸芴甲酯(OPA-FMOC)为衍生剂进行柱前衍生[17],根据峰面积和浓度之间的关系绘制氨基酸标准曲线,采用相同方法检测培养基中相对应的20 种氨基酸峰面积,根据标准曲线计算各氨基酸的浓度。
2.3 统计学分析
应用SPSS 统计分析软件对实验结果进行统计学分析,数据以平均值加减标准差的形式表示,p < 0.05表示存在显著性差异。
3 结 果
3.1 NK-92 细胞在培养过程中的氨基酸消耗分析
NK-92 细胞以 1×105cells·mL-1的密度接种于添加 2 mmol·L-1谷氨酰胺的 SFM-MIN 培养基培养 2 d 后检测培养基中20 种氨基酸浓度,计算培养2 d 后各氨基酸浓度变化差值如图1 所示。由图1 可知,NK-92细胞对不同氨基酸的需求存在较大的差异。天冬氨酸、谷氨酸、甘氨酸、丙氨酸和脯氨酸浓度呈升高趋势,其余氨基酸浓度都有不同程度的降低,谷氨酰胺、丝氨酸、亮氨酸、异亮氨酸和苏氨酸这5 种氨基酸浓度变化幅度较大,其中以谷氨酰胺变化量最为显著。进一步分析上述消耗较大的氨基酸的代谢特点,发现丝氨酸、苏氨酸亮氨酸和异亮氨酸合成的原料来均与糖代谢过程中的丙酮酸有关。由此可推测,NK-92 细胞对谷氨酰胺和丙酮酸两种营养物质有较高的需求。
3.2 谷氨酰胺和丙酮酸钠最适添加浓度的确定
使用 minitab 软件设计了两因子四水平正交实验,其中X 代表培养基中谷氨酰胺浓度,浓度分别为0.0、 6.5、13.0、19.5 mmol·L-1,Y 代表培养基中丙酮酸钠浓度,浓度为分别为 0.0、2.5、5.0、7.5 mmol·L-1(如表 1),共产生 16 个正交实验组。将 NK-92 细胞以 1×105cells·mL-1的密度接种于上述 16 种实验组无血清培养基中,每48 h 取样计数,并补充新鲜培养基和IL-2,调回最初接种密度,培养至10 d 时,测定各实验组的总细胞扩增倍数,结果如表2,图2 和表3 所示。

图1 NK-92 细胞在SFM-MIN 中培养2 d 的氨基酸浓度差(n = 3)Fig.1 Difference of amino acid concentration for NK-92 cells cultured in SFM-MIN for 2 days (n = 3)

表1 2 因子4 水平正交设计Table 1 Two-factor four-level orthogonal design
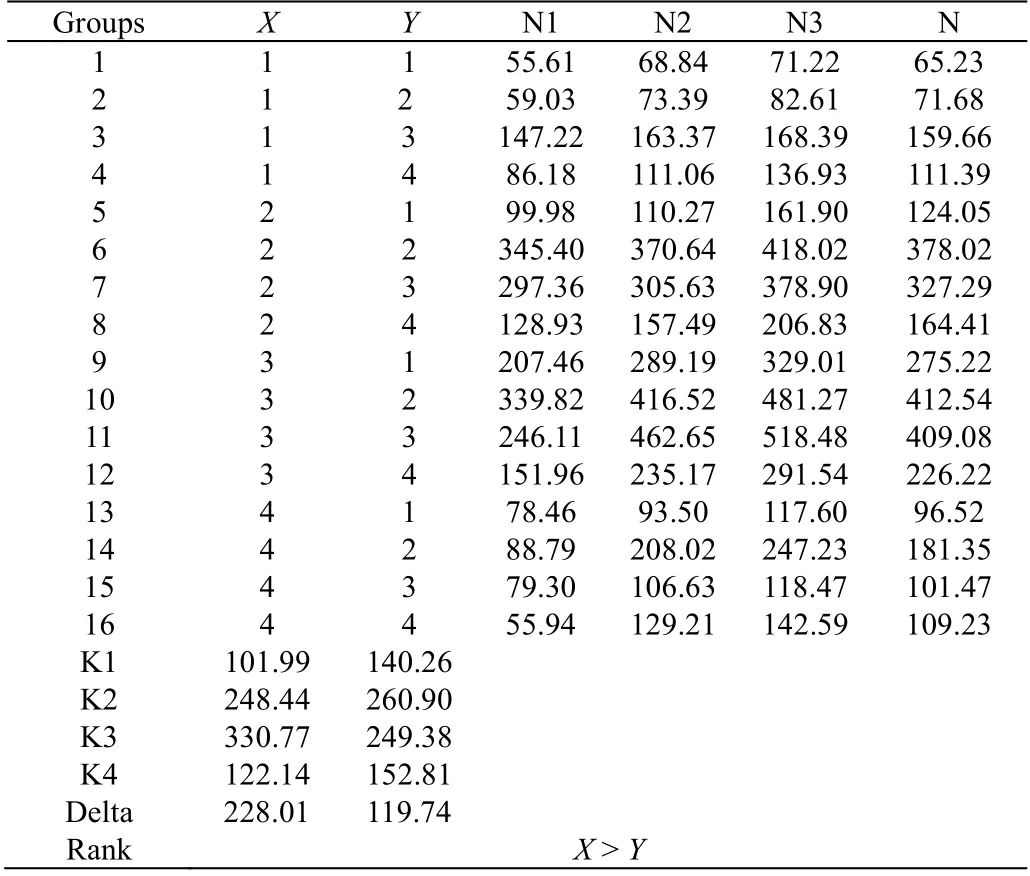
表2 正交实验结果(n = 3)Table 2 Results of the orthogonal experiments (n = 3)

图2 均值主效应图(n = 3)Fig.2 Main effect maps of mean values (n = 3)
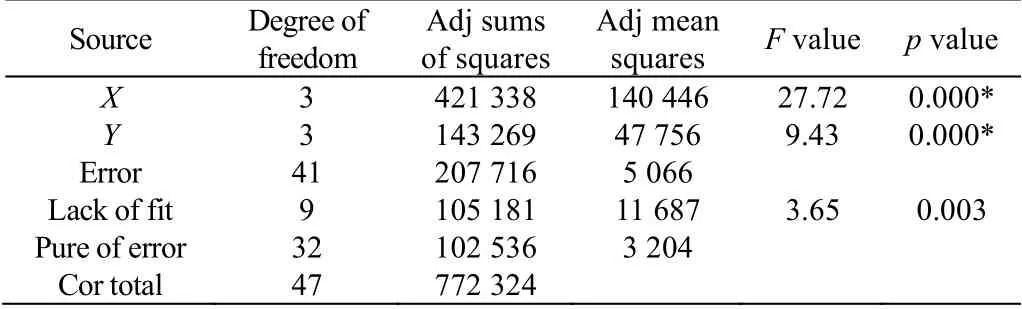
表3 正交实验结果方差分析Table 3 Analysis of variance of the orthogonal experiment results (*: p < 0.05)
N 为3 个平行实验组的总细胞扩增倍数N1~N3 的平均值,进一步求出X、Y 2 种因子对应4 种不同水平的N 值平均值,即K1、K2、K3、K4,得到均值主效应图即图2。由实验结果可知,在上述添加浓度范围中,谷氨酰胺最适水平为13.0 mmol·L-1,丙酮酸钠最适水平为2.5 mmol·L-1。
将正交实验的数据进行方差分析,结果如表3 所示,谷氨酰胺和丙酮酸钠对总细胞扩增倍数都有显著性的影响(p < 0.05)。
3.3 谷氨酰胺和丙酮酸钠促进未冻存NK-92 细胞的扩增效率
将提高了谷氨酰胺和丙酮酸钠浓度至上述最适水平的商业化α-MEM 有血清培养基和无血清培养基T009 为实验组,不额外添加谷氨酰胺和丙酮酸钠的原始培养基为对照组,以细胞活性、总细胞扩增倍数、培养物中CD3-CD56+细胞比例和细胞杀伤活性为指标,评价提高谷氨酰胺和丙酮酸钠浓度对NK-92 细胞扩增的影响。
图3为NK-92细胞在提高了谷氨酰胺和丙酮酸钠浓度的有血清培养基中的扩增情况。从图3(A)可知,培养过程中实验组NK-92 细胞活性与对照组相当;培养15 d 后,总细胞扩增倍数为(3 712.85±225.74) 倍,显著高于对照组的(2 255.93±243.00) 倍 (p < 0.05) (图 3(B));培养物中 CD3-CD56+细胞比例为(89.63±1.56)%,与对照组中 CD3-CD56+细胞比例(88.37±2.74)% 相近(图 3(C))。
图4 为NK-92 细胞在提高了谷氨酰胺和丙酮酸钠浓度的无血清培养基中的扩增情况。从图4(A)中可见,在培养前期,两种培养基中的NK-92 细胞活性相当,但培养到第9 d,对照组中NK-92 细胞活性开始降低,而实验组中NK-92 细胞的活率可以稳定维持在90% 以上;到第11 d 时,细胞活性已显著高于对照组 (p < 0.05);培养 15 d 的总细胞扩增为(3 193.59±199.99)倍,显著高于对照组的(1 917.16±242.87) 倍(p < 0.05) (图4(B));培养物中CD3-CD56+细胞比例为(90.37±0.90)%,显著高于对照组中CD3-CD56+细胞比例(82.68±2.28)% (p < 0.05) (图4(C))。可见,提高培养基丙酮酸钠和谷氨酰胺浓度促进了NK-92细胞的扩增。

以效靶比10:1 将扩增15 d 的NK-92 细胞与K562 共培养,分别评价在提高了谷氨酰胺和丙酮酸钠浓度的有/无血清培养基中扩增后的NK-92 细胞的杀伤功能,结果如图5 所示。由图可知,在有/无血清实验组中培养的NK-92 细胞的杀伤活性均值分别为63.53% 和71.22%,高于对照组57.23% 和62.63%。可见,提高培养基中丙酮酸钠和谷氨酰胺浓度提高了NK-92 细胞的杀伤活性。
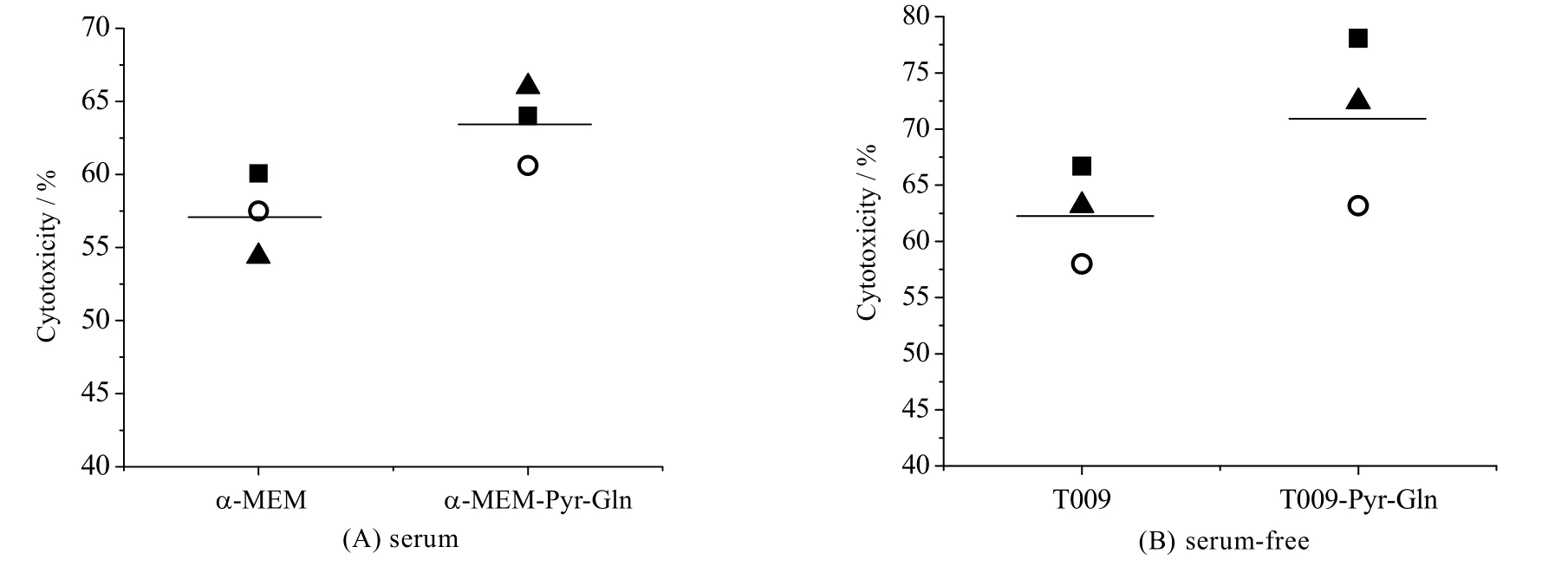
图5 有血清和无血清培养基中谷氨酰胺和丙酮酸钠对NK-92 细胞杀伤活性的影响(n = 3)Fig.5 Effects of glutamine and sodium pyruvate on the cytotoxicity of NK-92 cells in serum and serum-free media (n = 3) *: p < 0.05
3.4 谷氨酰胺和丙酮酸钠促进冻存后NK-92 细胞扩增效率
进一步将冻存7 d 后复苏的NK-92 细胞以5×105cells·mL-1的密度接种于有/无血清培养基中,以谷氨酰胺和丙酮酸钠浓度提高至上述最适水平为实验组,不额外添加谷氨酰胺和丙酮酸钠的原始培养基为对照组,以细胞活性、总细胞扩增倍数、培养物中CD3-CD56+细胞比例、胞内ATP 含量和细胞杀伤活性为指标,评价提高谷氨酰胺和丙酮酸钠浓度对冻存后NK-92 细胞生长的影响。
培养过程中的细胞活性、总细胞扩增倍数和培养物中 CD3-CD56+细胞比例如图 6 所示。由图 6(A)可知,在有血清实验组培养基中培养冻存复苏后的NK-92 细胞,培养1 d 后,其细胞活性可以快速由73.90%提高到90% 以上,而对照组中NK-92 细胞活性在第4 d 才上升至90%;且在培养的1~7 d,实验组中细胞的活性均显著高于对照组 (p < 0.05);由图6(B)可见,在无血清实验组培养基中,培养1 d 后,其细胞活性可以快速由73.90% 提高到80% 以上,而对照组中NK-92 细胞活性在第3 d 才上升至80% 以上;且在培养的1~5 d,其活性均显著高于对照组 (p < 0.05);在有/无血清实验组培养基中,培养15 d 的总细胞扩增分为 (3 117.74±120.21) 倍和 (1 282.35±162.68) 倍,显著高于对照组的 (1 031.86±111.13) 倍和(672.06±68.18)倍 (p < 0.05) (图6(C)~(D));同时,在有血清培养基中,培养物中CD3-CD56+细胞比例为(91.53±2.14)%,与对照组中CD3-CD56+细胞比例(89.67±1.44)% 相近(图6(E))。而在无血清培养基中,培养物中CD3-CD56+细胞比例为(92.06±2.28)%,显著高于对照组中CD3-CD56+细胞比例(84.81±1.74)%(p < 0.05) (图6(F))。可见,提高丙酮酸钠和谷氨酰胺浓度促进了冻存复苏后NK-92 细胞的扩增。

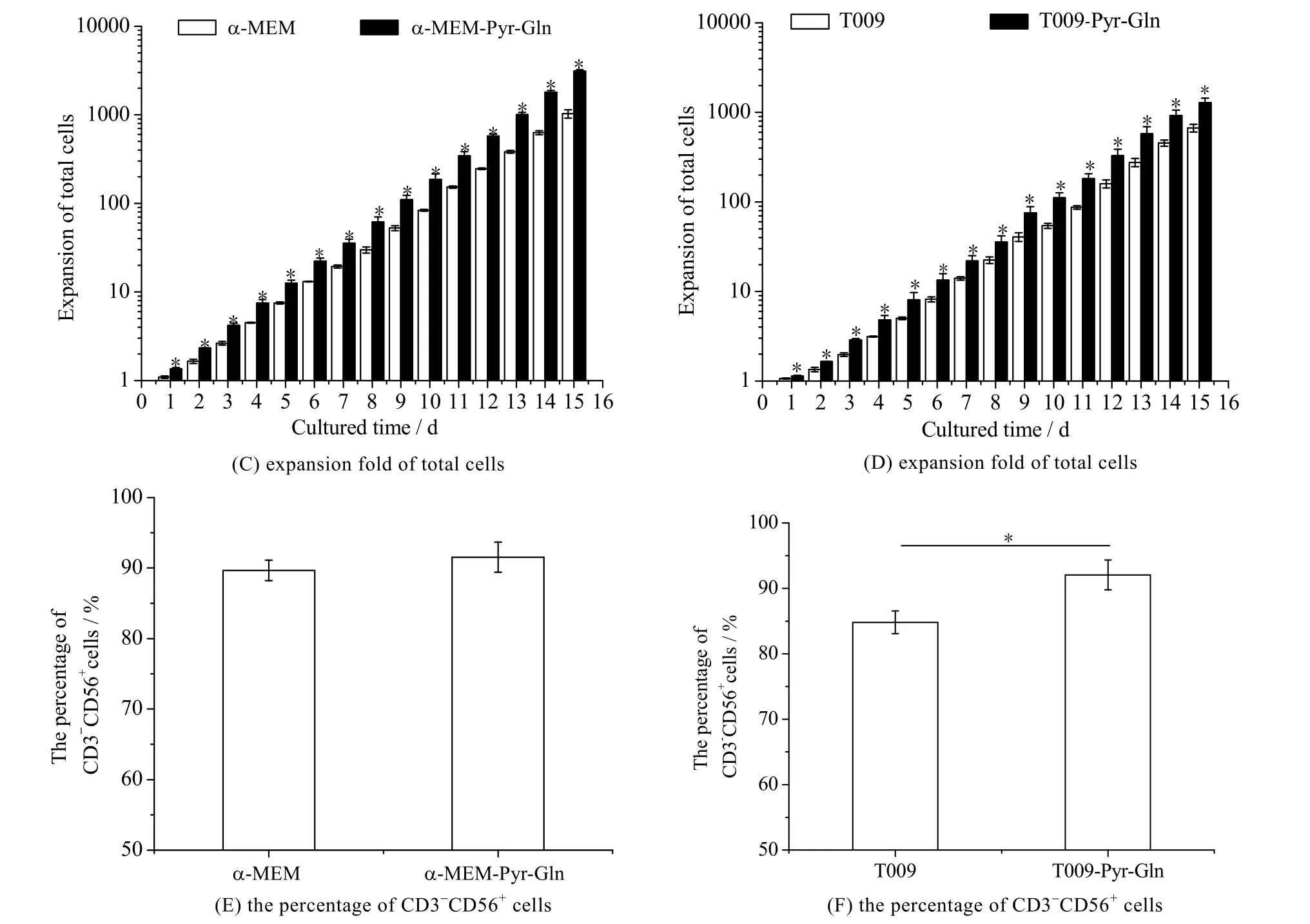
图6 有/无血清培养基中谷氨酰胺和丙酮酸钠对冻存后NK-92 细胞活性、总细胞扩增倍数和CD3-CD56+ 细胞比例的影响(n = 3)Fig.6 Effects of glutamine and sodium pyruvate on NK-92 cell activity, expansion fold of total cells and the percentage of CD3-CD56+ cells inserum/serum-free media after cryopreservation (n = 3) *: p < 0.05
谷氨酰胺和丙酮酸钠与细胞的能量代谢密切相关,ATP 是细胞内微环境的重要组成,是细胞合成代谢的关键因子,因此进一步分析NK-92 细胞在提高了谷氨酰胺和丙酮酸钠浓度的有/无血清培养基中培养1~4 d 的胞内ATP 水平,结果如图7 所示。由图可知,冻存后NK-92 细胞胞内ATP 都呈现先上升后下降的变化趋势,并都高于冻存复苏起始值;培养1 d 后,NK-92 细胞胞内ATP 均迅速回升,但在有/无血清培养基实验组中扩增的细胞胞内ATP 水平显著高于对照组(p < 0.05);而在培养第3 d,其细胞胞内ATP 水平显著低于对照组(p < 0.05)。由于ATP 作为细胞的“能量通货”,其胞内水平是生成和消耗共同作用的结果。提高谷氨酰胺和丙酮酸钠浓度有助于三羧酸循环的转运,从而促进胞内ATP的生成。在培养早期细胞扩增不明显,细胞胞内ATP 生成大于消耗,使得培养早期胞内ATP 水平高,有助于细胞快速进入扩增状态;而后期细胞进入快速扩增状态,需要更多的ATP 支持其快速生长,因而胞内ATP 水平低。由此可见,谷氨酰胺和丙酮酸钠的补充提升了细胞的能量代谢水平从而促进了复苏后NK-92 细胞的体外扩增。

图7 有血清和无血清培养基中谷氨酰胺和丙酮酸钠对冻存后NK-92 细胞胞内ATP 的影响(n = 3)Fig.7 Effects of glutamine and sodium pyruvate on intracellular ATP of NK-92 cells in serum and serum-free media after cryopreservation (n = 3) *: p < 0.05
进一步评价扩增后的NK-92 细胞的功能,结果如图8 所示。由图可知,在有/无血清实验组中培养冻存复苏的NK-92 细胞的杀伤活性均值分别为62.33% 和64.63%,高于对照组54.65% 和56.70%。可见,补充谷氨酰胺和丙酮酸钠提高了NK-92 细胞的杀伤活性。

图8 在有血清和无血清培养基中谷氨酰胺和丙酮酸钠对冻存后NK-92 细胞杀伤活性的影响(n = 3)Fig.8 Effects of glutamine and sodium pyruvate on the cytotoxicity of NK-92 cells after cryopreservation in serum and serum-free medium (n = 3) *: p < 0.05
4 结 论
本文基于自主设计的成分简单、明确的SFM-MIN 无血清培养基分析了NK-92 细胞氨基酸代谢特性,发现谷氨酰胺、丝氨酸、亮氨酸、异亮氨酸和苏氨酸这5 中氨基酸在培养过程中消耗最为显著,其中尤以谷氨酰胺为甚。考虑苏氨酸,丝氨酸、亮氨酸以及异亮氨酸其合成途径均与丙酮酸有关,因此确定谷氨酰胺和丙酮酸钠为关键营养组分,通过正交实验确定了其最适浓度分别为13.0 和2.5 mmol·L-1,并将含12.5% 胎牛血清和12.5% 马血清的α-MEM 培养基和市售的商业化无血清培养基T009 中谷氨酰胺和丙酮酸钠的浓度至提高至最适浓度,提高了未冻存和冻存后NK-92 细胞的扩增倍数和杀伤活性。该结果为免疫细胞快速、高效扩增提供思路。
