基于主要表型性状的不同采集地斑茅种质资源遗传多样性分析
2020-08-04王春芳余兴华王先宏杨清辉
王春芳 余兴华 王先宏 杨清辉

摘 要:为提高斑茅种质资源利用效率,本研究对来自不同地区的76份斑茅种质资源的株高、茎径、节间长、叶长、叶宽、锤度6个表型性状进行遗传多样性分析。结果表明,来自不同地区的斑茅,6个表型性状平均变异系数在14.6%~53.4%之间,其中株高和节间长的变异系数相对较高,分别为53.4%和37.7%;锤度和叶长的变异系数相对较低,分别为19.1%和14.6%。株高、茎径、叶宽、叶长、节间长之间表现为极显著正相关,其中株高与节间长的相关系数最大,为0.818;锤度与株高、节间长呈负相关,相关系数分别为?0.232和?0.263。主成分分析表明,前2个主成分方差累计贡献率高达72.58%,其中第1主成分方差贡献率达到51.21%,第2主成分贡献率为21.37%。采用聚类分析方法,不同的种质资源被分为8大类群,第Ⅴ和Ⅵ类群的种质综合性状表现最好,这2个类群是优良的斑茅种质资源,可在甘蔗育种中加以挖掘利用。
关键词:甘蔗;斑茅;表型性状;遗传多样性中图分类号:S31 文獻标识码:A
Genetic Diversity Analysis Based on Main Phenotypic Characteristics ofErianthus arundinaceumGermplasm from Different Regions
WANG Chunfang1, YU Xinghua2, WANG Xianhong1*, YANG Qinghui1*
1. College of Agronomy and Biotechnology, Yunnan Agricultural University, Kunming, Yunnan 650201, China; 2. Wenshan Academy of Agricultural Sciences, Wenshan, Yunnan 663099, China
Abstract: The germplasm resource, genetic diversity including plant height, stem diameter, internode length, leaf length, leaf width and brix of 76 clones ofE. arundinaceum, which were collected from different regions was analyzed. The average coefficient of variation of the six phenotypic traits was 14.6%–53.4%, the coefficient of variation of plant height and internode length was 53.4% and 37.7%, respectively, and the coefficient of variation of brix and leaf length was 19.1% and 14.6%, correspondingly. There was a significant positive correlation among plant height, stem diameter, leaf width, leaf length and internode length, among which the correlation coefficient between plant height and internode length was 0.818. Meanwhile, brix was negatively correlated with plant height and internode length, with correlation coefficients of ?0.232 and ?0.263, respectively. Principal component analysis showed that the cumulative contribution rate of the first two principal components was 72.58%, of which the contribution rate of the first principal component was 51.21%, and that of the second principal component was 21.37%. By cluster analysis, 76 clones were divided into eight groups and the comprehensive characters of V and VI groups were the best. The two groups are excellent E. arundinaceumgermplasm resources, which could be excavated and utilized in sugarcane breeding program.
Keywords: Saccharum spp.;Erianthus arundinaceum; phenotypic characteristics; genetic diversity
2.2 种质资源表型性状间的相关分析
对76份斑茅种质资源的6个主要农艺性状进行相关性分析(表4),结果表明,株高与茎径、叶宽、节间长呈极显著正相关,相关系数分别为0.477、0.619和0.818;茎径与叶长、叶宽、节间长呈极显著正相关,相关系数分别为0.56、0.774、0.522;叶宽与节间长呈极显著正相关关系,相关系数为0.556;锤度和株高、节间长呈负相关,相关系数分别为?0.232和?0.263。由此可知,当斑茅茎径越粗壮、叶片越宽、节间长越长,其生长越高大、越茂盛;株高越高,节间长越长,其锤度越小。
2.3 种质资源表型性状的主成分分析
为了综合评价各因素对斑茅种质资源遗传变异的主导作用,以斑茅种质资源6个表型性状的数据为基础,计算各表型性状的主成分载荷和贡献,结果见表5。前5个主成分方差累计贡献率
高达98.41%,第1主成分贡献率为51.21%,株高、茎径、叶宽和节间长为决定因子,表明第1主成分是反映植株株型的主要因子,但锤度为负向指标,载荷值为?0.194,表明在挖掘利用斑茅种质资源时,在株型和糖分含量的选择上需要取舍。第2主成分贡献率为21.37%,节间长和株高为主要正向指标,载荷值分别为0.231、0.442,表明第2主成分主要反映植株株高的因子;而叶长和茎径为主要的负向指标,载荷值分别为?0.746、?0.335,表明叶子的长短、茎径的粗细与植株的高矮也需要在斑茅种质资源的利用过程中进行权衡。第3主成分贡献率为14.99%,其中锤度的载荷值最大,为0.932,因此第3主成分主要是斑茅种质资源糖分含量的决定因子。第4、5和6主成分分别在节间长、茎径和株高上具有最大的载荷绝对值,分别为0.614、0.703、0.693。综上表明,斑茅种质资源在植株的高矮、叶片的形态以及锤度均具有较大的遗传多样性。
由表5可知,第1主成分和第2主成分的特征值大于1,分别为1.753、1.132。以第1主成分为横坐标,第2主成分为纵坐标,根据84份种质资源的来源和种属进行分组,76份斑茅种质资源6个表型性状的数据为基础,进行主成分分析作图(图1)。分别由割手密、五节芒、河八王和蔗茅种质资源各2份组成的外群(OG)距离其他的斑茅资源分组较远,说明其遗传关系较远,差异最大。来源于缅甸(MMR)、西藏(XZ)和中国云南(YN)的分组距离相对较近,且聚为一大组,主要表现为植株高度、葉片宽度和节间长3个性状的相关性。其余的6个分组,包括福建和江西组成的分组(FJ&JX)、广东和广西组成的分组(GD&GX)、海南(HN)、MG分组(包括河南、湖北、湖南和陕西)、四川(SC)、美国(US),聚为另一大组。另外,福建和江西组成的分组(FJ&JX)与云南(YN)的分组距离最远,说明其遗传关系最远。综上所述,从6个表型性状的数据进行主成分分析的结果基本符合各种质资源的地域分布特性,地理分布较远的斑茅资源,遗传关系上也较远。
2.4 不同斑茅种质资源的聚类分析
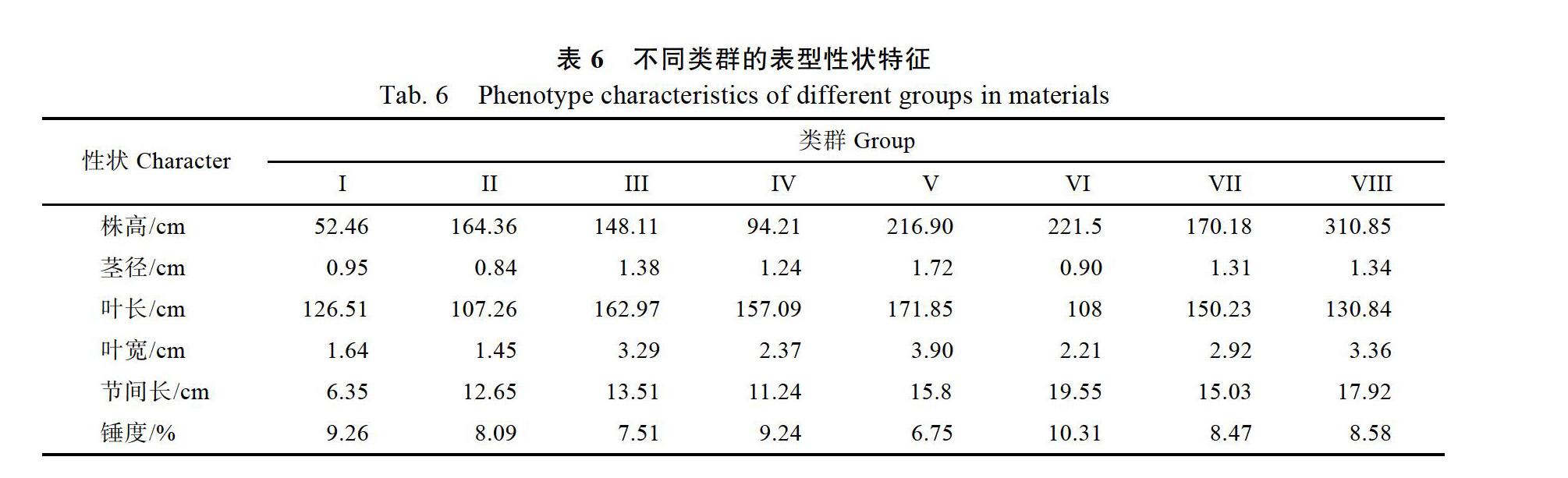
聚类分析既可以揭示类群间的遗传差异与相互关系,又可以了解类群内不同无性系间的遗传相似性。76份斑茅种质资源的6个表型性状数据标准化处理后,根据欧氏距离进行聚类分析。在欧氏距离约为100处,可以把全部76份种质资源划分为8个类群(图2),其中的外群主要被聚类在第Ⅱ和Ⅵ类群中。不同类型的表型性状特征见表6。
第Ⅰ类群包括16份斑茅种质,其表现为植株矮小,节间长最小;第Ⅱ类群包括6份斑茅种质,该类群斑茅种质的茎径最细,叶片宽度最窄;第Ⅲ类群包括7份种质,第Ⅵ类群包括17份种质,这两个类群植株的6个表型性状表现适中;第Ⅴ类群包括4份种质,该类群斑茅种质的茎径最粗,叶片最大,但锤度最低,可能是具有高光效潜能的优良种质资源;第Ⅵ类群包括3份斑茅种质,该类材料节间长最长、锤度最高,但叶片长度较短,可以作为甘蔗品种改良的优质资源;第Ⅶ类群包括4份斑茅种质,各性状指标表现适中;第Ⅶ类群包括19份斑茅种质,为最大的一个类群,西藏地区一半的斑茅种质被聚于该类群,最突出的特点是植株高大,各表型性状表现均较好,在甘蔗育种中可以作为优良种质加以利用。
3 讨论
3.1 斑茅种质资源的表型性状差异
本研究根据对来自不同采集地的斑茅种质资源的表型性状分析表明,76份斑茅种质资源的6个表型性状差异较大,其株高变化范围为30.6~ 441.2 cm,茎径变化范围为0.63~1.786 cm,叶长变化范围为94.4~184.4 cm,叶宽变化范围为0.44~ 4.72 cm,节间长变化范围为2.78~29.2 cm,锤度变化范围为5%~12%。而前人普遍认为斑茅植株高大,株高为2~6 m,叶片长度1~2 m,叶片宽度2~5 cm[30-31]。本研究拓宽了斑茅种质资源的表型变化范围,除了生长茂盛、高大的斑茅种质,也存在一些矮小的斑茅种质资源,为甘蔗育种提供了更广泛的种质资源。
3.2 不同采集地斑茅种质资源的遗传多样性
斑茅近年来越来越受到国内外甘蔗育种界的重视,具有丰富的遗传变异。前人研究表明,斑茅种质资源的形态特征存在广泛的变异,各形态性状之间显著相关,聚类结果与形态相似性有关[20]。另外,斑茅的表型特征具有较高的遗传变异,平均变异系数为14.95%~32.15%,遗传距离和斑茅的不同采集区域存在一定的相关性[21]。在本研究中,6个表型性状的平均变异系数变化范围为14.6%~53.4%,除锤度以外,各表型性状之间表现出显著的相关性,各种质资源的遗传关系也表现出地域分布特性,这些结果都与前人的研究结果相一致[18, 22]。聚类分析在揭示类群间的遗传差异和类群内不同材料的遗传相似性上方便直观。本研究对不同采集地的斑茅种质资源进行聚类分析表明,76份种质被分为8个类群,聚类结果同样与形态相似性有关。前人对来自广西的50份斑茅资源的4个表型性状进行分析,变异系数范围为12.71%~16.14%[32],本研究中来自广西的3份斑茅种质的平均变异系数高达43.6%,主要表现在株高的变化上,其株高变化范围为35.4~ 317.4 cm,表明广西斑茅资源具有较丰富的遗传多样性。
3.3不同采集地斑茅种质资源的应用潜力
斑茅丰富的遗传多样性为优质资源的挖掘和利用提供了更多的可能[33-34],特别来自不同地区、具有不同优良性状的种质资源其遗传多样性更高。在供试材料中,来自西藏地区的18份斑茅种质,大部分株高相对较高,叶片相对较长、较宽,节间长相对较长,其中有一半的种质聚类到第Ⅷ类群,这些斑茅种质各性状表现优异,可作为优质资源加以利用,特别是在高光效、高产品种改良中可以发挥重要作用。来自江西地区的6份材料植株相对矮小,节间长较短,可以为矮化育种提供基因资源。来自海南、广西、广东、福建地区的19份斑茅资源表型变化相对较大,植株形态各异,可以为挖掘利用特殊优异资源提供更大的可能。
本研究主要基于斑茅种质资源的6个主要表型性状进行遗传多样性的初步分析,由于表型性状容易受到自然环境改变的影响,以及群体样本量少,并不能系统和准确地评估种质资源的遗传关系[35-36]。因此,应结合现代生物学技术对种质资源的遗传多样性进行系统分析评价,利用分子遗传标记以及具体的生理生化指标对斑茅种质资源进行分析,为挖掘利用斑茅优良种质资源提供理论依据。
参考文献
[1] 李奇偉. 现代甘蔗品种改良技术[M]. 广州: 华南理工大学出版社, 2003.
[2] 彭绍光. 国内外甘蔗种质资源的采集保育与利用研究概述[J]. 西南农业学报, 1990(3): 90-96.
[3] 刘少谋. 几个斑茅后代作为甘蔗杂交亲本的利用效果[J]. 甘蔗糖业, 1992(3): 1-6.
[4] 王丽萍, 蔡 青, 范源洪, 等. 甘蔗(Saccharum)与斑茅(Erianthus arundinaceus)远缘杂交利用研究[J]. 西南农业学报, 2007(4): 721-726.
[5] 文明富, 杨俊贤, 潘方胤, 等. 甘蔗遗传改良研究进展[J]. 广东农业科学, 2016, 43(6): 58-63.
[6] 周耀辉. 斑茅分类地位的研究[J]. 甘蔗糖业, 1989(6): 4-6, 17.
[7] 蔡 青, 范源洪, Aitken K, 等. 利用AFLP进行“甘蔗属复合体”系统演化和亲缘关系研究[J]. 作物学报, 2005(5): 551-559.
[8] 何顺长. 云南省甘蔗野生资源植物开发利用前景的探讨[J]. 云南农业大学学报, 1987(1): 105-111.
[9] 周耀辉, 黄鸿能, 殷培军, 等. 海南甘蔗育种场甘蔗种质种性研究(Ⅱ)[J]. 甘蔗糖业, 1997(3): 2-7.
[10] 沈万宽. 斑茅的杂交利用价值探讨[J]. 甘蔗, 2002(3): 1-5.
[11] Tagane S, Tagane M Y, Ponragdee W,et al. Cytological stu?dy ofErianthus procerusandE. arundinaceus(Gramineae) in Thailand[J]. Cytologia, 2011, 76(2): 171-175.
[12] 廖兆周, 劳方业, 周耀辉, 等. 具有斑茅种质的耐旱甘蔗品系的选育[J]. 作物学报, 2002(6): 841-846.
[13] 周 珊, 高轶静, 张保青, 等. 斑茅割手密复合体杂交利用过程野生特异基因遗传分析[J]. 植物遗传资源学报, 2019, 20(3): 718-727.
[14] 杨清辉, 李富生, 肖凤回, 等. 斑茅染色体和植物学性状观察研究[J]. 云南农业大学学报, 1997 (4): 28-31.
[15] 萧凤迴, 扬清辉, 周彩萍, 等. 云南斑茅的染色体和植物学性状观察初报[J]. 云南农业大学学报, 1992(1): 59-63.
[16] 梁绪振, 鄢家俊, 白史且, 等. 斑茅种质资源形态性状的变异研究[J]. 草业科学, 2011, 28(7): 1307-1314.
[17] 刀志学, 鄢家俊, 张建波, 等. 斑茅野生种质资源的考察与收集[J]. 植物遗传资源学报, 2013, 14(5): 816-820
[18] 徐超华, 陆 鑫, 马 丽, 等. 斑茅种质资源的表型性状及遗传多样性[J]. 湖南农业大学学报(自然科学版), 2014, 40(2): 117-121.
[19] 刘 浩, 周闲容, 于晓娜, 等. 作物种质资源品质性状鉴定评价现状与展望[J]. 植物遗传资源学报, 2014, 15(1): 215-221.
[20] 梁绪振. 斑茅种质资源遗传多样性及生物质能潜力评估[D]. 雅安: 四川农业大学, 2011.
[21] 徐超华, 陆 鑫, 刘新龙, 等. 甘蔗近缘种蔗茅Erianthus fulvus考察收集与表型性状初步研究[J]. 植物遗传资源学报, 2014, 15(5): 962-966.
[22] 刘 洋, 刘新龙, 苏火生, 等. 海南甘蔗野生种资源的收集与遗传多样性初析[J]. 中国农学通报, 2013, 29(1): 199-208.
[23] 胡存脈, 王先宏, 余兴华, 等. 藏南甘蔗野生种质资源的考察与植物学性状分析[J]. 云南农业大学学报(自然科学版), 2015, 30(3): 351-356.
[24] 中华人民共和国农业部. 农作物种质资源鉴定技术规程 甘蔗: NY/T 1488—2007 [S]. 北京: 中国农业出版社, 2008: 1-4.
[25] 陆 鑫, 毛 钧, 应雄美, 等. 甘蔗创新种质的因子分析与聚类分析[J]. 西南农业学报, 2011, 24(6): 2072-2076.
[26] 赵铭钦, 王玉胜, 刘国顺, 等. SPSS软件在烤烟品种综合评价中的应用[J]. 中国农学通报, 2006(10): 128-130.
[27] 赵 勇, 赵培方, 胡 鑫, 等. 基于农艺性状分级对317份甘蔗种质资源的评价[J]. 中国农业科学, 2019, 52(4): 602-615.
[28] 张革民, 杨荣仲, 刘海斌, 等. 割手密主要数量性状的主成分及聚类分析[J]. 西南农业学报, 2006(6): 1127-1131.
[29] 刘新龙, 马 丽, 蔡 青, 等. 云南甘蔗品种表型性状的遗传多样性分析[J]. 植物遗传资源学报, 2010, 11(6): 703-708.
[30] 陈守良. 中国植物志[M]. 北京: 科学出版社, 1997, 10(2): 39-44.
[31] 程天聪. 云南甘蔗种质资源考察及有性杂交育种研究[J]. 西南农业学报, 1992(1): 14-19.
[32] Liu X H, Song H Z, Zhang G M,et al. Phenotypic variation and genetic diversity in the collections ofErianthus Arundinaceus(Retz.) Jesw.[J]. Sugar Tech, 2017, 19(4): 359-367.
[33] 陈勇生, 陈西文, 邓海华, 等. 斑茅蔗BC1后代产量与品质的通径分析[J]. 甘蔗糖业, 2011(2): 1-4.
[34] 谢 静, 吴建涛, 符 成, 等. 甘蔗与斑茅远缘杂交不同世代材料光合特性分析[J]. 分子植物育种, 2018, 16(8): 2723-2729.
[35] 杨 继. 植物种内形态变异的机制及其研究方法[J]. 武汉植物学研究, 1991(2): 185-195.
[36] 黄玉新, 段维兴, 张保青, 等. 138份国外引进甘蔗品种(系)宿根性评价[J]. 云南农业大学学报(自然科学版), 2019, 34(4): 564-570.
