氮添加对长序榆C、N、P养分含量及非结构性碳水化合物含量的影响*
2020-07-28洪琮浩雷小华汪俊峰闫道良
洪琮浩 洪 震 雷小华 汪俊峰 闫道良
(1.北京林业大学生物科学与技术学院 北京 100083; 2. 华东药用植物园科研管理中心 丽水 323000; 3. 浙江丽水市莲都区城区林业工作中心站 丽水323000; 4. 浙江农林大学林业与生物技术学院 杭州 311300)
氮是植物的生命元素,影响着植物的生长、营养元素的吸收利用、光合生理以及能量物质的贮藏等。工农业的快速发展以及自然环境的变化,导致氮(N)沉降持续增加(Reayetal., 2008),引起土壤酸化、森林衰退和物种多样性改变等生态环境问题(段娜等, 2019; Sparriusetal., 2012),严重影响陆地生态系统的生产力和稳定性(苏洁琼等, 2012; 何玉惠等, 2015)。在植物生长受氮限制的区域,氮沉降会增加土壤无机氮,有益于植物的生长(孙宇等, 2019; 陈磊等, 2018);而氮相对丰富的区域,氮沉降却不能发挥氮素的营养作用,反而会抑制植物的生长,降低植物的生产力(Luetal., 2014; 赵俊晔等, 2006)。氮施加试验也表明,适量添加氮会促进植物的生长、生物量的增加和净光合速率(Pn)的提高,从而显著提高植物的生产能力(周晓兵等, 2010; 张云海等, 2013)。不同的植物对氮肥的需求量有差异,且对氮肥添加的响应机制各异,不同的植物对氮添加的响应并不一致。
长序榆(Ulmuselongata)属榆科(Ulmaceae)榆属(Ulmus),是濒临灭绝的极小种群野生植物,亟需拯救保护(国家林业局&农业部, 1999; 2011),主要零星分布在福建、江西、浙江和安徽的中低海拔常绿阔叶林地区。近年来在长序榆的分布区如江西的一些林区随降雨输入的氮约为60 kg·hm-2a-1,而在浙江金华则为17.2~21.3 kg·hm-2a-1,氮沉降所导致的林区土壤氮含量的增加远远高于森林对氮的需求(5~8 kg·hm-2a-1)(李欠欠等, 2010)。因此,在氮沉降持续增加的大背景下,探讨濒危植物长序榆对氮沉降的响应就十分必要。本文从模拟氮沉降的角度,研究氮添加对长序榆主要养分元素氮(N)、磷(P)积累及其之间的计量关系和非结构性碳水化合物含量的影响,以期为长序榆的濒危机制的研究探寻新的思路,为更好地拯救长序榆提供参考。
1 材料与方法

1.2 试验方法 小心抖落根部基质,充分洗净后,把根、茎、叶分开,置于烘箱中烘干至恒质量,各部分粉碎过筛备用。采用重铬酸钾氧化-外加热法测定植物样品中全碳含量,硫酸-双氧水消解,半微量凯氏定氮法测定全氮含量,钼锑抗比色法测定全磷含量; 采用硫酸蒽酮法测定可溶性总糖含量,淀粉含量测定采用酶水解法(张韫, 2011; 张志良等, 2005),非结构性碳水化合物(NSC)含量为可溶性糖与淀粉含量之和。用Li-3100A叶面积仪 (Li-Cor, Lincoln, Nebraska, USA) 测量叶面积并计算比叶面积(specific leaf area, SLA),其计算公式为比叶面积=叶面积/叶片干质量(cm2·mg-1)。
利用Li-6400便携式光合测定系统(LI-COR, Inc., Lincoln, USA)测定叶片气体交换参数。选取叶位相同生长状况一致的叶片,连续3天对同一叶片进行指标测定,测定时间为上午9:00—10:00,叶室温度设定为25 ℃,测定期间的外界气温为(24~26 ℃),叶室CO2浓度约为400 μmolCO2·mol-1,叶室气体流速为500 μmol·s-1,光合有效辐射(PAR)为720 μmol·m-2s-1,与长序榆生长环境光照强度基本一致。测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、气孔限制值(Ls)等叶片气体交换参数。每个处理测定10枚叶片,每次测定重复5次,并计算植物叶片的水分利用效率(WUE)=Pn/Tr。
1.3 数据处理 利用SPSS 22.0 软件对数据进行统计分析,采用Duncan法比较各处理间差异性,采用Excel 2013作图。
2 结果与分析
2.1 氮添加对长序榆生长的影响 不同施氮水平处理下,长序榆的根、茎生物量并没有呈现明显的差异,而高氮处理(N4)明显降低了长序榆叶片干物质的积累,相比对照降低28.28%(图1)。比叶面积是植物碳摄取策略的关键叶性状之一,高氮处理(N4)明显降低了比叶面积(图2),由此说明长序榆在高氮处理下植物光捕获能力呈现显著下降。
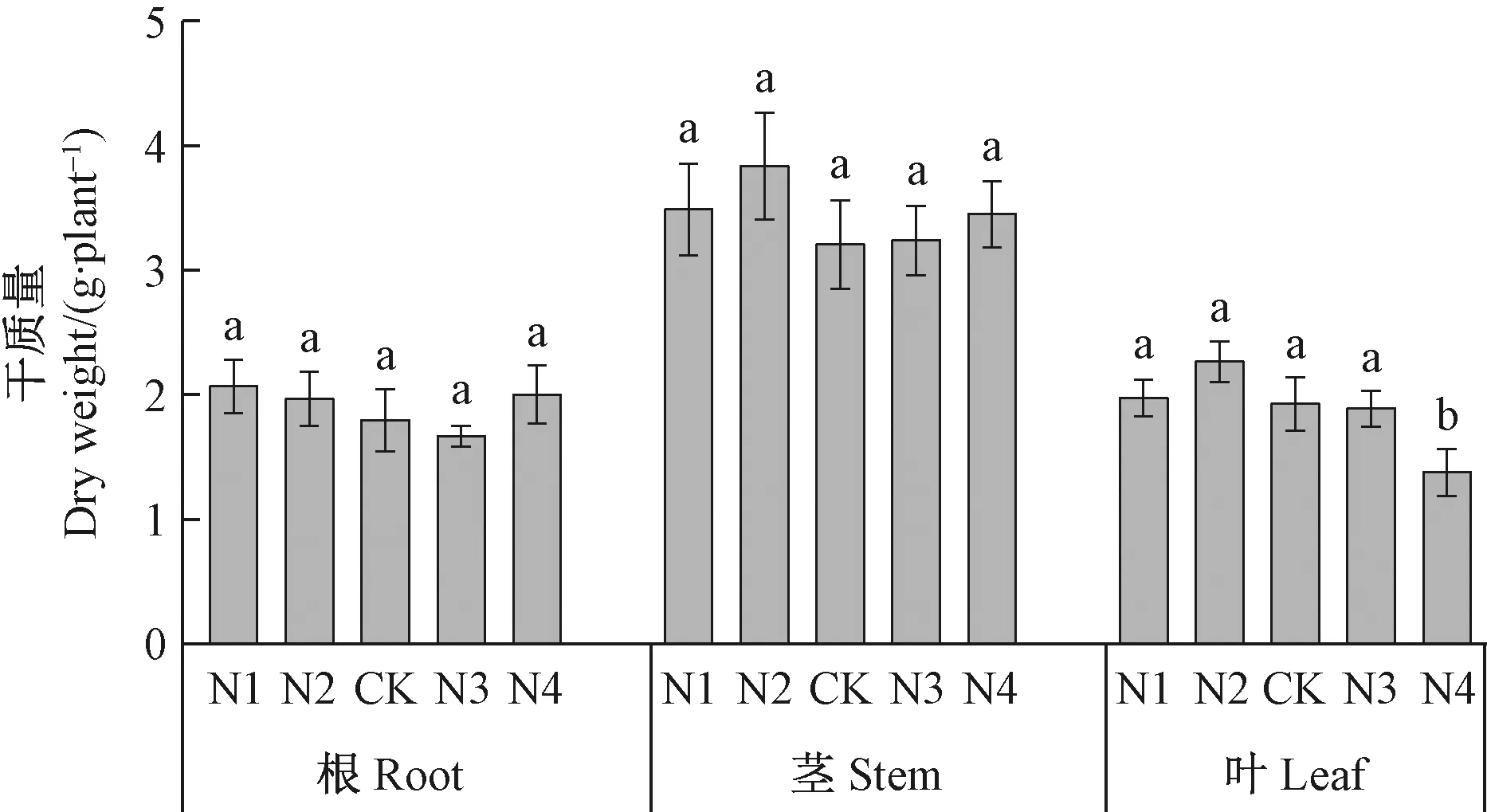
图1 氮添加对长序榆干物质积累的影响Fig.1 Effects of nitrogen addition on Dry matter accumulation in U. elongata
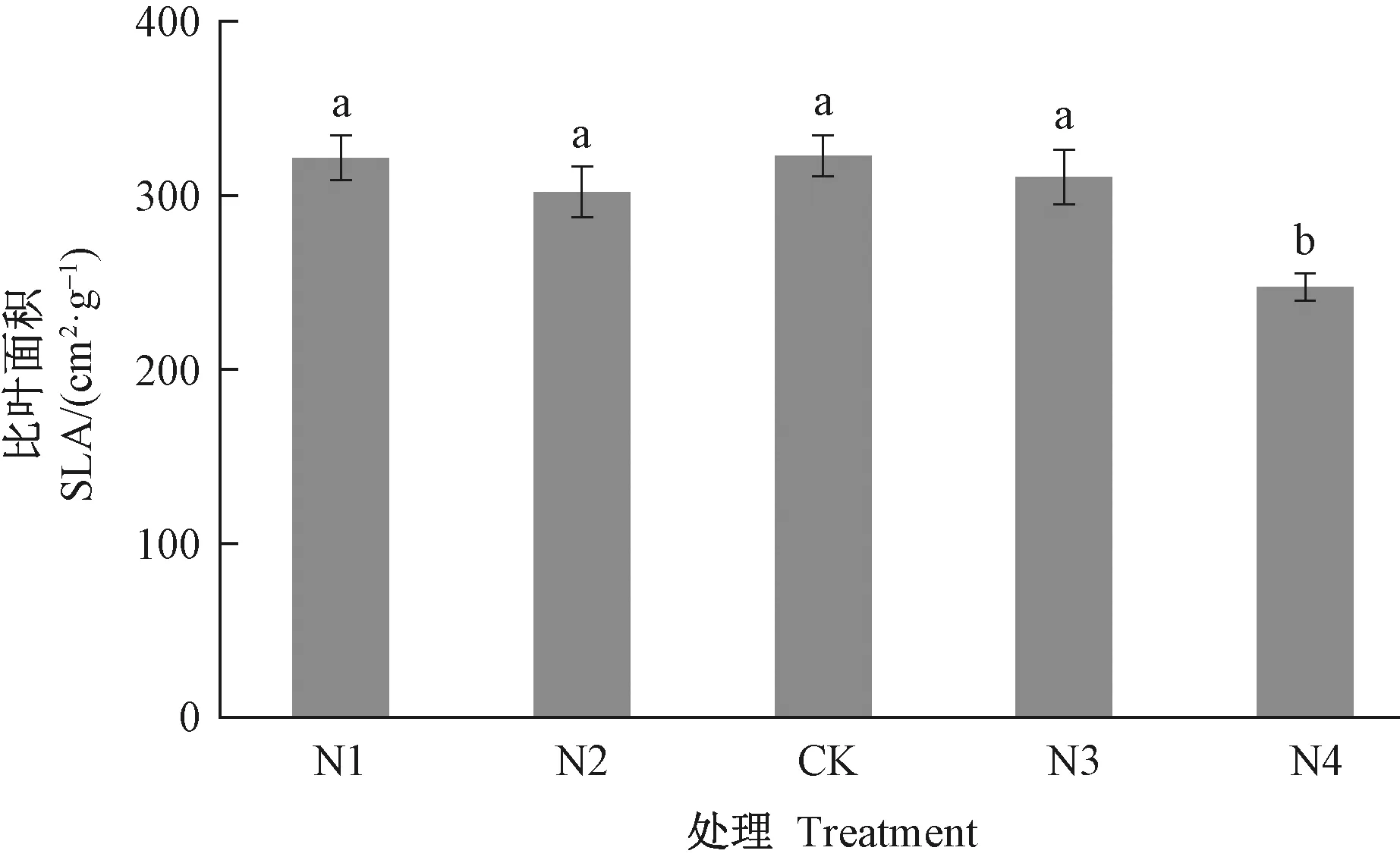
图2 氮添加对长序榆比叶面积的影响Fig.2 Effects of nitrogen addition on specific leaf area of U. elongata
2.2 氮添加对长序榆C、N、P含量及其生态化学计量特征的影响 与对照相比,不同施氮水平下,低氮和高氮均会提高根中总碳含量。茎中的总碳含量表现各异,低氮处理显著促进了茎中碳的积累,N1处理下茎中总碳的含量是对照的1.17倍。当氮浓度在CK~N4范围内,茎中的氮含量呈现相对稳定的状态。低氮(N1)和高氮(N3、N4)均会降低叶中碳的积累,与对照相比差异显著(P<0.05)(图3)。

图3 氮添加对长序榆总碳含量的影响Fig.3 Effects of nitrogen addition on total carbon content ofU. elongata
CK处理下,长序榆根、茎、叶中的总氮含量均相应明显低于低氮和高氮处理下的根、茎、叶中总氮含量(图4)。在高氮处理下,根、茎、叶中的总氮含量随着施氮水平的增加而显著增加,N4处理下长序榆根、茎、叶中总氮含量分别是对照的1.73、2.15和3.08倍。当施氮量在N1~CK范围内时随着施氮量的降低,根、叶中的氮含量明显增加。长序榆植株中的磷素含量则随着施氮水平的增加,根、茎、叶中总磷含量均显著下降(图5)。N1处理下根、茎、叶中总磷含量分别是N4处理的2.35、2.39、2.70倍。

图5 氮添加对长序榆总磷含量的影响Fig.5 Effect of nitrogen addition on total phosphorus content of U. elongata
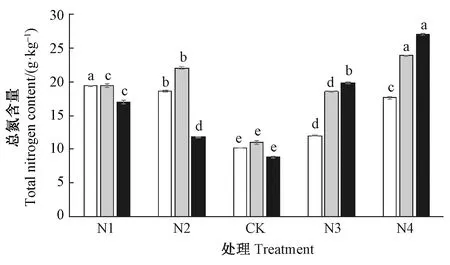
图4 氮添加对长序榆总氮含量的影响Fig.4 Effects of nitrogen addition on total nitrogen content of U. elongata
根、茎、叶中的C∶N呈现大致相同的变化趋势,低氮和高氮均降低了C∶N比。CK处理的C∶N比显著高于低氮和高氮处理。C∶P和N∶P均表现为高氮(N4)处理最高(表1)。

表1 氮添加对长序榆C、N、P计量学的影响①Tab.1 Effects of nitrogen addition on chemometrics of C, N and P in U. elongata
2.3 氮添加对长序榆NSC含量的影响 根和茎中的可溶性糖和淀粉含量随施氮水平的变化呈现相似的变化趋势(图6、图7),低氮(N1)和高氮(N4)均会提高可溶性糖和淀粉含量。N2处理下,根和茎中的可溶性糖含量最低,淀粉含量则是CK处理最低(P<0.05)。高氮处理(N3和N4)显著降低叶中可溶性糖和淀粉含量。NSC含量随施氮水平的增加,在根和茎中先降低后升高,在叶中为先升高后降低(图8),与可溶性糖和淀粉含量在根、茎和叶中的变化趋势相似。

图8 氮添加对长序榆NSC含量的影响Fig.8 Effects of nitrogen addition on NSC content in U. elongata

图6 氮添加对长序榆可溶性总糖含量的影响Fig.6 Effects of nitrogen addition on soluble total sugar content in U. elongata

图7 氮添加对长序榆淀粉含量的影响Fig.7 Effect of nitrogen addition on starch content of U. elongata
2.4 长序榆根、茎、叶中NSC分配格局 长序榆不同器官NSC含量存在明显差异(表2)。可溶性糖含量高低分布顺序为叶 > 根和茎; 淀粉含量高低分布顺序为根和茎 > 叶,NSC含量高低分布顺序是根 > 叶和茎。
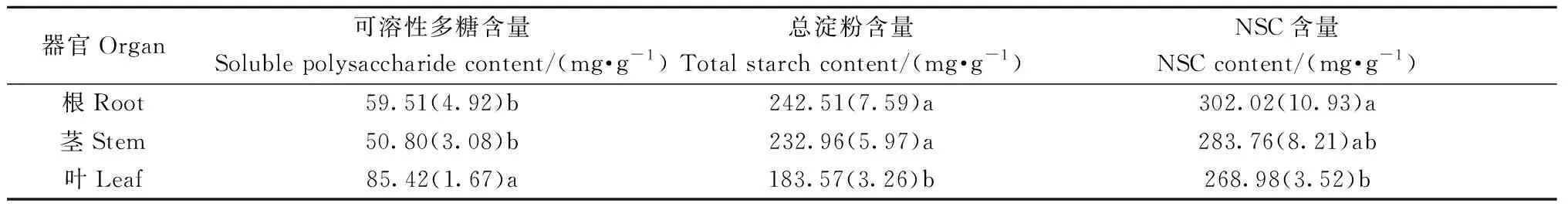
表2 非结构性碳水化合物在根、茎、叶中分配格局①Tab.2 Distribution pattern of non-structural carbohydrate in roots, stems and leaves of U. elongata
2.5 氮添加对长序榆光合性能的影响 由表3可知,高氮(N4)导致Pn、Gs、Tr、WUE和Ls不同程度下降,Ci表现上升。与对照相比,Pn、Gs、Tr、WUE和Ls分别下降93.54%、71.43%、76.13%、72.73%和44.83%,而Ci上升25.74%。低氮(N1)处理并没有明显影响长序榆的Pn、Gs、Tr、WUE和Ls。

表3 氮添加对长序榆光合性能的影响Tab.3 Effects of nitrogen addition on photosynthetic performance of U. elongata
3 讨论
3.1 氮添加对长序榆总碳含量、氮磷养分及其化学计量学特征的影响 随着施氮量的增加,长序榆根中的总碳含量表现先降低后升高,茎中是先降低后保持相对稳定,而叶中则表现先升高后降低。长序榆根、茎、叶中总碳积累对氮添加的响应并不一致,高氮能够促进叶中的碳向根中转移。叶片和根是源库关系,叶片中的总碳含量高,有利于光合产物向根中转移,由此增加根中能量物质的贮藏,提高长序榆对逆境的适应。当外源氮超过一定阈值(CK浓度)时,长序榆根、茎、叶中的氮含量才随着外源氮添加的增加而升高。当外源氮浓度低于CK时,根、茎、叶中的氮随着氮添加的增加而降低,这可能是因为该浓度范围的氮促进源、库器官更快地生长,从而“稀释”植株根、茎、叶中氮的浓度。随着氮添加量的增加,长序榆植株内的P含量显著下降。研究表明,生态系统内长期氮增加,会降低系统内磷循环率,加剧土壤受磷限制(Wangetal., 2014),从而严重制约植株对磷的吸收。磷与植株对逆境的抗性密切相关,施用磷肥可以增加植株的抗旱性,增强对矿质元素的吸收及干物质的积累(张小明等, 2016),因此,氮添加抑制长序榆植株内磷的积累,会严重影响其对环境干扰的抵抗力,加速其老化死亡。这与王乔姝怡等(2016)的研究中N添加没有明显改变乔木层植物和灌木层植物叶片磷含量的结论不一致,但与氮添加显著减少了苔藓植物叶片磷含量的结论相一致。
在植物与环境相互作用的过程中,保持能量平衡和化学元素平衡是维持生态系统稳定性的重要前提,其中碳∶氮∶磷化学计量学的特征是各种生态过程研究的核心内容。研究表明,持续氮添加所引起的植物营养元素的失衡,会改变植物群落物种的组成,进而影响群落物种N∶P的值。当持续氮沉降所引起的土壤有效氮增加时,生长缓慢的物种会被高效利用氮的物种所取代,进而改变物种的优势度,尤其对渐危的极小种群物种来说,面临被排挤的风险(Nordinetal., 2005)。对长序榆氮添加研究表明,高氮(N4)显著提高了根、茎、叶N∶P值,由此会导致对P吸收的限制,造成长序榆P胁迫。同时,研究发现高氮引起C∶P显著增加,这可能与P的相对匮乏有密切关系。高氮并没有使C∶N值提高,与相对适宜的施氮量CK比较,却显著低于CK处理的C∶N值。以上结果表明,高施氮量并没有提高长序榆对氮的利用效率,反而降低其对氮的利用效率,因而增加了长序榆在群落中被高效利用氮素的物种所代替的危机。
3.2 氮添加对长序榆非结构性碳水化合物(NSC)含量及光合参数的影响 长序榆根、茎、叶中NSC含量对氮添加响应的规律不同。根、茎中可溶性糖和淀粉含量随着氮添加的增多均表现先降低后升高的趋势。根、茎中可溶性多糖及NSC含量均在高氮(N4)处理下最高,淀粉含量则在N3处理下最高; 叶中的可溶性多糖、淀粉和NSC含量分别在N1、N2和N2处理下最高。综上所述,较高浓度的氮添加能够促进长序榆根、茎中非结构性碳水化合物的积累,叶中则是在低氮下有较多的积累,说明高氮促进长序榆叶中(源)的NSC组分向根、茎中(库)转移,从长序榆能量代谢的角度来说,这种高氮处理下能量转移的策略,可能更有利于长序榆对高氮处理下所引起的逆境环境的适应。
氮是限制植物光合作用的重要因素,在一定氮添加范围内光合性能随施氮量的增加而提高(Nakajietal., 2002; 陈翠莲等, 2019)。本研究同时测定了长序榆在不同氮处理下的光合性能,结果表明,高氮(N4)显著降低了长序榆的Pn、Gs、Tr、WUE和Ls,提高了Ci,从而抑制长序榆的光合性能,影响光合物质的积累,最终会削弱其在生态系统的竞争能力,加剧其濒危的程度。这可能是由于高氮下,光合作用的关键酶Rubisco酶合成受阻,从而抑制长序榆的光合性能,也可能是由于高氮使叶片中的N∶P值显著增加,体内矿质营养失衡所致。
本研究仅对模拟氮添加的大背景下,长序榆的氮磷养分积累、非结构性碳水化合物含量变化及光合指标进行分析,而关于不同种类的施肥组合以及与土壤水分的交互作用对长序榆生长乃至在群落中竞争趋势的研究仍需要通过进一步的试验验证。
4 结论
高氮处理(262.3 mg·L-1)会使长序榆体内积累过多的氮,磷含量显著下降,N∶P值升高,从而加剧植株磷限制。同时,叶中碳水化合物含量降低,而根、茎中则会积累更多的碳水化合物。长序榆把更多能量物质储存在根、茎中的生存策略,以应对高氮的不利影响。因此,在长序榆的栽培管理中,可考虑合理控施氮肥,以达到调节植株的N、P养分平衡,促进叶片光合性能,从而实现对长序榆的有效保护。
