黔西北岩溶区华山松(Pinus armandii)种群结构及动态特征
2020-06-29陈群利薛晓辉李望军
何 斌,李 青,陈群利,薛晓辉,李望军
(贵州省普通高等学校生物资源开发与生态修复特色重点实验室/ 生态工程学院,贵州工程应用技术学院,贵州 毕节 551700)
植物种群结构是植物个体生存能力与环境相互作用结果的体现[1],是一种动态平衡关系[2]。种群结构和动态特征是种群生态学研究的核心内容之一[3],可以反映种群对环境的适应能力[4],对阐明生态系统的维持、群落结构特征与演替规律、种群的生存现状和更新策略具有重要意义[5]。种群的年龄结构、静态生命表、生存分析和时间序列分析等是种群统计学的重要手段,不仅可以描述种群的生存现状,还能够为种群的发展趋势提供重要信息[6-7],近年来被广泛应用于植物种群的研究上[8-9]。
华山松(Pinusarmandii)属松科(Pinaceae)松属(Pinus)常绿乔木,为我国特有树种,分布范围广,在山西、陕西、贵州、云南等13个省份海拔1 000~3 300 m范围内均有分布,生长迅速、耐寒力强,能适应多种土壤,具有很强的水源涵养和水土保持等生态功能。华山松在中国西南喀斯特地区森林生态系统中亦占据重要地位,贵州西北部的喀斯特地区也是其分布区之一。同常态地貌上的森林植被相比,喀斯特森林是一类特殊的森林系统,在森林群落特征、演替更新以及种群与环境之间的相互作用等方面均有显著差异[10]。目前,对华山松群落的研究主要集中在植物区系分析、群落结构、物种多样性、优势树种生态位、生物量、养分循环等方面[11-16],但是,对喀斯特地区华山松种群结构与动态方面的研究尚鲜见报道。
鉴于此,笔者以黔西北华山松种群为研究对象,通过对其种群结构特征、静态生命表、存活曲线、生存分析和时间序列分析等方面的研究,试图探明华山松种群的生存现状、数量动态及发展趋势,为该物种的保护及资源管理提供科学依据,且对喀斯特森林生态系统结构和功能的维持以及退化森林生态系统的恢复重建具有重要意义。
1 研究区概况与研究方法
1.1 研究区概况
草海国家级自然保护区位于贵州省威宁县西南侧,地处云贵高原乌蒙山区腹地,地理坐标介于26°47′35″~26°52′10″ N,104°09′23″~104°20′10″ E之间。研究区属亚热带高原季风气候区,具有日照丰富、冬暖夏凉、冬干夏湿的气候特征;年平均气温 10.9 ℃,最热月(7 月)平均气温 17.3 ℃,最冷月(1月)平均气温 2.1 ℃;年均降水量 903.6 mm,主要集中在 5—8 月,占全年降水总量的70.4%;年平均蒸发量948.7 mm,主要集中在3—5 月,占全年蒸发总量的43.9%;年平均日照时数 1 455.5 h,无霜期平均 190.5 d。土壤主要为黄棕壤,此外还有棕色石灰土、石质土、沼泽土等。研究区地带性植被为中亚热带半湿润常绿阔叶林,主要由常绿栎、栗、胡桃和枫杨等阔叶树组成,受全球气候变化和人类活动的影响,逐渐逆向演替为目前的次生针叶林和针阔混交林,主要由华山松、云南松(Pinusyunnanensis)、滇杨(Populusyunnanensis)和灌丛等多种植被类型组成。
华山松群落为人工飞播造林形成的纯林,乔木层郁闭度约75%,树高平均为17 m,伴生有槲栎(Quercusaliena)、杉木(Cunninghamialanceolata)和云南松;灌木层覆盖度约30%,主要有滇榛(Corylusyunnanensis)、贵州小檗(Berberiscavaleriei)、火棘(Pyracanthafortuneana)、扁刺峨眉蔷薇(Rosaomeiensis)、灰叶栒子(Cotoneasterhebephyllus)等;草本层覆盖度约20%,主要有蛇莓(Duchesneaindica)、鱼腥草(Houttuyniacordata)、一年蓬(Erigeronannuus)、车前草(Plantagoasiatica)、老鹳草(Geraniumwilfordii)、猪殃殃(Galiumaparine)、珠光香青(Anaphalismargaritacea)、车轴草(Galiumodoratum)、紫花地丁(Violaphilippica)等。
1.2 研究方法
1.2.1样地设置与调查方法
2017年7—8月,在全面踏查的基础上,根据种群分布范围及生境片断化状况,在草海国家级自然保护区内设置15块20 m×30 m的样地,采用GPS测定各样地的地理坐标和海拔,同时记录坡度、坡向等基本信息。对样地内所有乔木进行每木检尺,测量胸径、树高、冠幅等;在样地内按梅花形分别设置5块2 m×2 m的灌木样方和1 m×1 m的草本样方,记录种名、数量、高度和盖度等。
1.2.2年龄结构划分
一般采用生长锥钻取木芯的方法来确定树种年龄[17],但是由于实际工作中无法追踪研究区内所有华山松个体的生长周期,利用生长锥取木芯确定其个体的真实年龄较为困难,且缺乏研究区内华山松的解析资料,以及考虑到自然保护区内对树种的严格保护要求,故该研究不采用钻取木芯的方法来确定树木年龄。由于多数乔木树种形成层有规律的分生,使其胸径与树龄之间存在一定的线性关系,大多数学者采用“空间代替时间的方法”,即用径级结构代替年龄级结构。根据华山松的生长规律,基于野外样地调查数据,将植株胸径(DBH)从小到大看作时间顺序,参照文献[18-19]将华山松种群划分为9个级别:Ⅰ级(DBH<3 cm)、Ⅱ级(3 cm≤DBH<6 cm)、Ⅲ级(6 cm≤DBH<9 cm)、Ⅳ级(9 cm≤DBH<12 cm)、Ⅴ级(12 cm≤DBH<15 cm)、Ⅵ级(15 cm≤DBH<18 cm)、Ⅶ级(18 cm≤DBH<21 cm)、Ⅷ级(21 cm≤DBH<24 cm)、Ⅸ级(DBH≥24 cm)。统计各龄级个体数,绘制华山松种群径级结构图。
1.2.3种群动态量化方法
华山松种群结构动态量化分析参考陈晓德[20]的方法,计算公式如下:
(1)种群内相邻两龄级间个体数量(Vn)的变化动态:
(1)
(2)忽略外部干扰的种群数量变化动态指数(Vpi):
(2)
(3)现实中,Vpi会受到外部干扰,还与年龄级数量(k)和各龄级内的个体数(Sn)有关, 可将式(2)修正为
(3)
(4)完全随机干扰下种群所承担的最大风险概率(P极大):
(4)
式(1)~(4)中,Vn为种群从n到n+1级的个体数量变化动态指数;Sn、Sn+1分别为第n和n+1级种群个体数量;Vpi为整个种群结构的数量变化动态指数;k为种群大小级数量。 当Vn、Vpi、Vpi′为正值时,说明种群个体数量呈增长趋势;当Vn、Vpi、Vpi′为0时,说明种群个体数量呈稳定的状态;当Vn、Vpi、Vpi′为负值时,说明种群个体数量呈衰退趋势[9]。
1.2.4种群静态生命表和存活曲线
参考BEGON等[21]静态生命表的编制方法,以径级代替龄级,编制华山松种群的静态生命表,主要包括以下参数:x龄级内现有个体数(Ax),x龄级开始时标准化存活个体数(lx),从x到x+1龄级间隔期内标准化死亡数(dx),从x到x+1龄级间隔期间死亡率(qx),从x到x+1龄级间隔期间还存活的个体数(Lx),从x龄级到超过x龄级的个体总数(Tx),进入x龄级个体的生命期望或平均期望寿命(ex),消失率(损失度)(Kx),具体计算公式如下:
lx=ax/a0×1 000,
(5)
dx=lx-lx+1,
(6)
qx=dx/lx×100% ,
(7)
Lx=(lx+lx+1) / 2 ,
(8)
(9)
ex=Tx/lx,
(10)
Kx=lnlx- lnlx+1。
(11)
式(5)~(11)中,ax为匀滑修正后x龄级内的存活个体数;a0为ax的初始值。
由于静态生命表是反映种群多个时代更迭动态过程的某一特定时间范围,并不能追踪种群的整个生活史[4],在编制过程中可能有死亡率为负的情况出现,这与静态生命表的数学假设不符[22],所以采用匀滑技术对调查数据进行处理[23]。以径级对应龄级为横坐标,标准化存活数的自然对数为纵坐标,绘制存活曲线。为了检验种群存活状况符合哪种Deevey曲线,运用HETT等[24]的检验方法对存活曲线进行拟合检验。
1.2.5生存分析
参考冯士雍[25]的计算方法,引入生存率函数S(i)、累积死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)4个函数分析华山松种群的动态变化规律,具体公式如下:
(12)
F(i)=1-S(i),
(13)
(14)
(15)
式(12)~(15)中,Si-1为i-1龄级存活率,%;Si为i龄级存活率,%;hi为i龄级宽度。
以径级对应龄级为横坐标,各生存函数值为纵坐标,分别绘制华山松种群的生存率曲线、累积死亡率曲线、死亡密度曲线和危险率曲线。
1.2.6时间序列预测
参考谢衷洁[26]的时间序列预测方法,采用一次移动平均法预测华山松种群未来经过2、4、6、8龄级后各龄级的个体数量。计算公式如下:
(16)
式(16)中,Mnt为未来经过n个龄级后t龄级种群的个体数量;n为需要预测的时间;t为龄级;Xk为k龄级内的个体数量。
2 结果与分析
2.1 华山松种群龄级结构
图1为华山松种群的龄级结构分布。随着龄级级数的增大,华山松种群个体数先增加后降低,在Ⅲ级达到最大。其中Ⅰ~Ⅱ龄级个体数占总数的26.47%,表明华山松种群幼苗和幼龄个体数不充足;Ⅲ~Ⅶ龄级的个体数最多,占总数的63.40%,表明华山松种群主要集中在此龄级范围;Ⅷ~Ⅸ龄级的个体数量最少,仅占总数的10.13%。综上所述,华山松种群整体表现出大、小龄级个体数少、中间龄级个体数多的纺锤型结构。
为了对华山松种群结构特征进行更准确的评价,采用种群动态量化方法分析其相邻龄级结构动态。相邻龄级间的数量动态指数(Vn)随龄级的增加波动较大,正值、0和负值均有出现。V1~V8的动态指数值分别为-20.00%、-3.23%、13.98%、6.25%、4.00%、5.56%、26.47%和76.00%,V1~V2值均小于0,说明Ⅰ、Ⅱ龄级的种群个体数量呈衰退动态关系;其他龄级的动态指数值均大于0,说明其他龄级的种群个体数量呈增长动态关系。不考虑外部干扰的情况下,种群数量变化动态指数(Vpi)为10.62;考虑外部干扰的情况下,种群数量变化动态指数(Vpi′)为0.10。与Vpi相比,Vpi′更趋近于0,说明华山松种群属于增长型种群,各龄级的个体数量在一定程度上影响了种群的年龄结构动态。随机干扰风险极大值(P极大)为0.93,说明华山松种群对外界的随机干扰敏感性较弱。
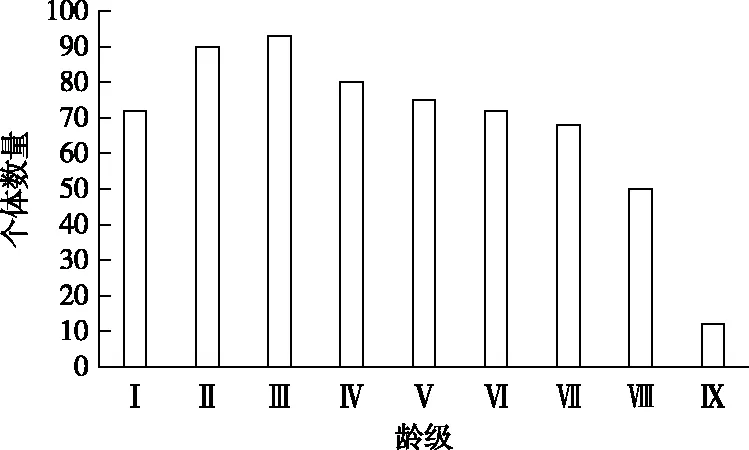
图1 华山松种群龄级结构
2.2 华山松种群静态生命表及存活曲线
从华山松种群静态生命表(表1)中可以看出,随着龄级增加,华山松种群各龄级的存活数逐渐减少,个体期望寿命(ex)逐渐降低,死亡率(qx)和消失率(Kx)逐渐增加,这与其生物学特性一致。这是因为幼龄林阶段种群个体树高、冠幅较小,群落内资源丰富,种群个体间竞争小,生存环境质量高,个体期望寿命高,死亡率和消失率低;随着林龄的增加,个体径级和冠幅不断增长,体积逐渐增大,需要占据更多的生存空间和养分,种内竞争加剧,群落内的资源无法完全满足所有个体的生长需要,同时个体达到一定年龄后生理机能衰退,个体期望寿命逐渐下降,死亡率和消失率逐渐升高。
存活曲线是借助于特定年龄存活的个体数量相对于时间作图得到的,反映了种群个体在各龄级的存活情况[19]。从华山松的存活曲线(图2)中可以看出,华山松种群的存活率整体呈下降趋势,第Ⅰ~Ⅴ龄级存活曲线较平缓,曲线斜率较小,个体存活数下降较慢;第Ⅴ~Ⅷ龄级存活曲线下降幅度增加,死亡率提高;第Ⅷ~Ⅸ龄级存活曲线的斜率最大,个体存活数下降最快。
按照Deevey存活曲线类型的划分标准,华山松种群的存活曲线介于Deevey-Ⅱ型和 Deevey-Ⅲ型之间。使用Hett和Loucks 的2种数学模型Nx=N0e-bx和Nx=N0x-b对华山松种群的存活曲线进行检验,建立如下模型:
y=8.199 9e-0.007 1x,R2= 0.713 3,
(17)
y=7.926 4x-0.224,R2= 0.496 7。
(18)
式(17)~(18)中,y为标准化存活数对数;x为龄级。
由于指数模型的R2值大于幂函数模型的R2值,因此华山松种群的存活曲线更趋近于Deevey-Ⅱ型。戈峰[27]认为Deevey-Ⅱ型又可以分为3个亚型:B1亚型,即不同时期种群个体成活量相差较大;B2亚型,即不同年龄段种群个体的死亡率基本相同;B3亚型,即种群幼年期死亡率较高,成年后的死亡率降低。对其进行直线拟合,建立如下模型:
y=-0.369 1x+7.726 3,R2=0.800 0。
(19)
结合图2可以看出,华山松种群的存活曲线属于Deevey-Ⅱ型中的B1亚型。
种群的死亡率和消失率曲线能够反映种群的动态变化过程。由华山松种群的死亡率和消失率曲线(图3)可知,华山松种群的死亡率和消失率曲线变化趋势基本一致,两者与龄级呈正相关关系,均随着龄级的增长而增加。这说明随着华山松的生长,个体逐渐进入乔木层,面临的生存竞争压力越来越大,同时由于乔木层个体间生态位重叠越来越严重,林内的生存空间和营养资源无法满足其生长需要,导致个体逐渐死亡、数量逐渐减少。
表1 华山松种群静态生命表
Table 1 Static life table ofP.armandiipopulation
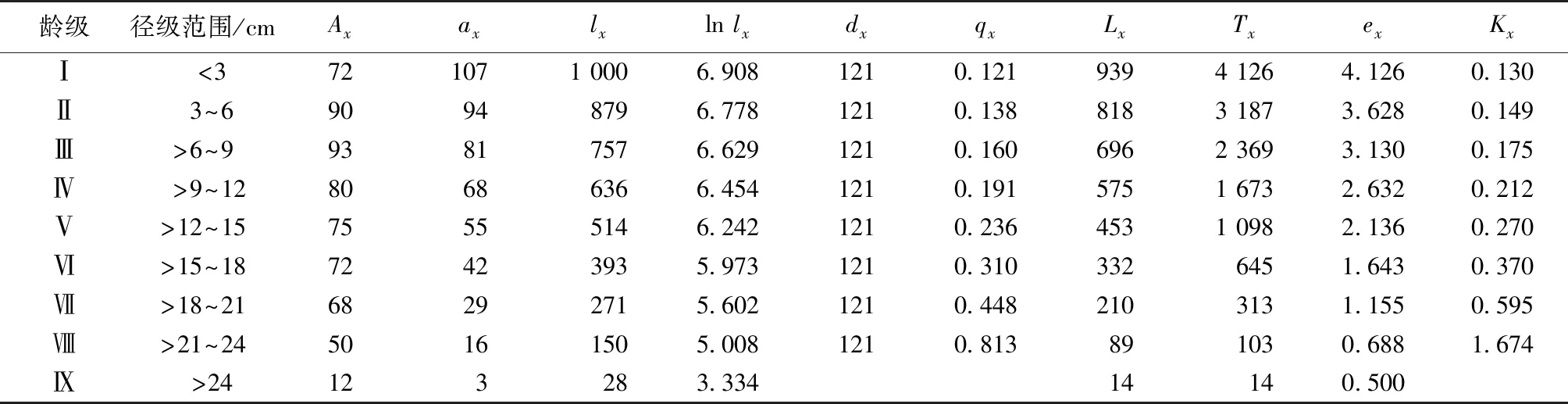
龄级 径级范围/cmAxaxlxln lxdxqxLxTxexKxⅠ<3721071 0006.9081210.1219394 1264.1260.130Ⅱ3~690948796.7781210.1388183 1873.6280.149Ⅲ>6~993817576.6291210.1606962 3693.1300.175Ⅳ>9~1280686366.4541210.1915751 6732.6320.212Ⅴ>12~1575555146.2421210.2364531 0982.1360.270Ⅵ>15~1872423935.9731210.3103326451.6430.370Ⅶ>18~2168292715.6021210.4482103131.1550.595Ⅷ>21~2450161505.0081210.813891030.6881.674Ⅸ>24123283.33414140.500
Ax为x龄级内现有个体数;ax为匀滑修正后x龄级内的存活个体数;lx为x龄级开始时标准化存活个体数;dx为从x到x+1 龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;Lx为 从x到x+1 龄级间隔期间还存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望或平均期望寿命;Kx为消失率。
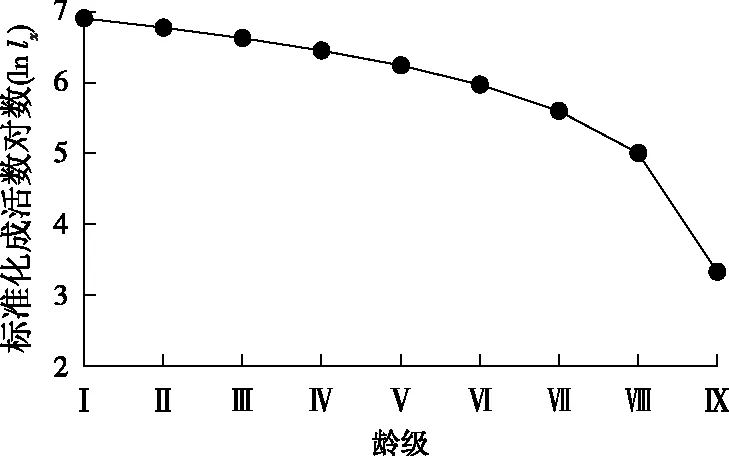
图2 华山松种群存活曲线Fig.2 Survival curve of P. armandii population
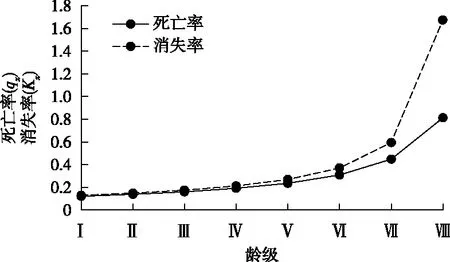
图3 华山松种群死亡率和消失率曲线
2.3 华山松种群生存分析
华山松种群生存分析结果(图4和图5)表明,生存率逐渐递减,累积死亡率逐渐递增,两者呈相反的变化趋势,符合物种的生物学特性;Ⅷ龄级后,生存率函数和累积死亡率函数变化均比较平缓,此时种群的生存率小于3%,累积死亡率大于97%,说明华山松种群Ⅷ龄级后逐渐走向生理枯竭。整体上看,死亡密度和危险率曲线两者反差明显,死亡密度曲线变化平缓,稳中有降,其值小于5.0%,表现为中幼龄死亡密度高于老林龄;危险率曲线呈逐渐增加的趋势,最高达66.7%。综合4 个生存函数的分析结果,发现华山松种群具有前期平稳、中期增长、后期衰退的特点。
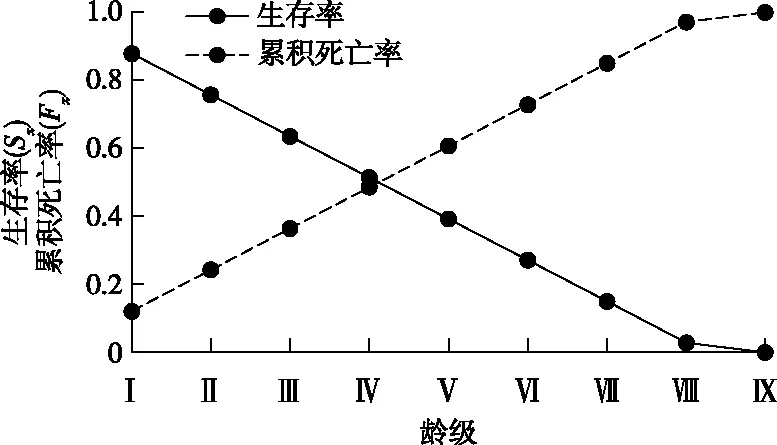
图4 华山松种群生存率和累计死亡率曲线Fig.4 Survival and cumulative mortality rates of P. armandii population
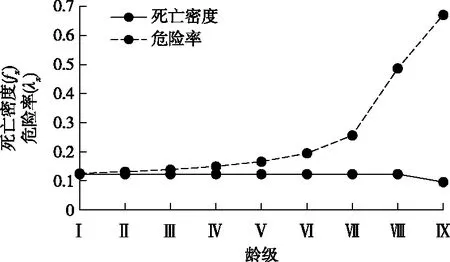
图5 华山松种群死亡密度和危险率曲线
2.4 华山松种群时间序列分析
根据华山松种群各龄级株树的统计结果,按照一次移动平均法预测未来2、4、6、8龄级后各龄级的个体数量(表2)。结果表明,未来2、4、6和8个龄级后,华山松种群个体数从Ⅴ龄级开始均有不同程度的增加,且龄级越高,未来经历的时间越长,种群个体数量增加幅度越大。以第Ⅸ龄级为例,在经历未来2、4、6、8龄级后分别增加了1.58、3.21、3.96和4.63倍,这表明华山松种群的增长主要依靠中、成林个体来维持,短期内会保持一定的增长趋势,但是该种群一旦达到生理年龄,必然会走向衰退。种群个体数在Ⅱ~Ⅲ龄级经过2个龄级后分别下降10.00%和1.07%,这表明华山松种群幼苗个体缺乏,不能保证幼、中龄阶段个体充分更新。综合其整体发展趋势来看,华山松种群难以保证其正常更新,如果不采取相应措施,未来种群会逐渐衰退。
表2 华山松种群动态变化的时间序列分析
Table 2 Time sequence analysis of age structure ofP.armandiipopulation
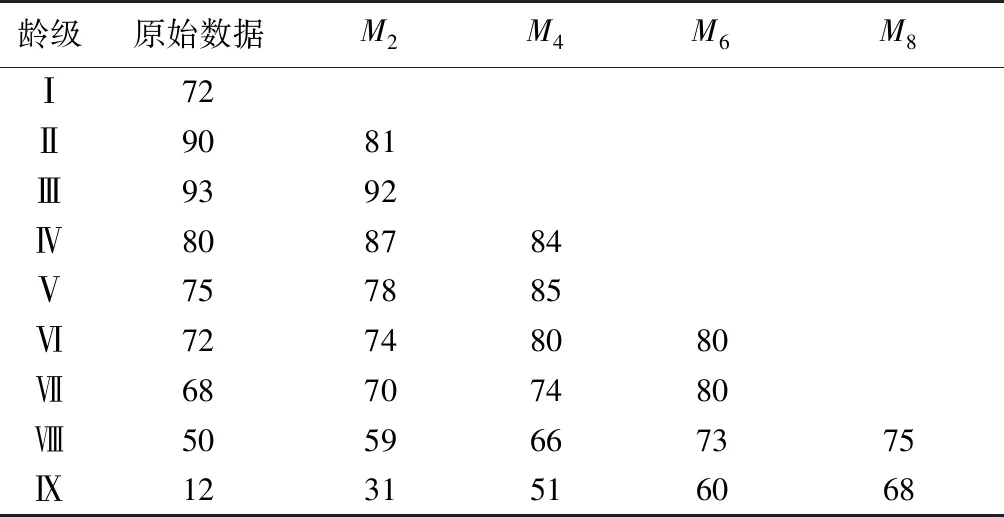
龄级原始数据M2M4M6M8Ⅰ72Ⅱ9081Ⅲ9392Ⅳ808784Ⅴ757885Ⅵ72748080Ⅶ68707480Ⅷ5059667375Ⅸ1231516068
M2、M4、M6和M8分别为经过 2、4、6和8 龄级时间后各龄级的种群大小。
3 讨论与结论
3.1 华山松种群的结构特征
植物种群数量动态可以反映种群不同阶段个体的组配情况,揭示种群的生存现状和更新策略[21]。植物种群结构是植物对环境条件适应性的一种体现,可以反映种群内部个体的生长发育过程[9]。从研究结果可知,华山松种群龄级结构呈两头小中间大的纺锤型结构,其中Ⅰ~Ⅱ龄级个体存活数占总数的26.47%,Ⅲ~Ⅶ龄级个体存活数占总数的63.40%,Ⅷ~Ⅸ龄级个体存活数占总数的10.13%,表明该种群为稳定型结构。这与数量动态指数反映出的种群结构存在一定差异。数量动态指数表明华山松种群动态存在一定的波动性,整体表现为增长型种群,但Ⅰ~Ⅱ龄级出现了负增长现象(Vi值小于0),这说明现有的华山松种群主要依靠成龄个体来维系其种群结构,种群的更新过程可能被延缓或者阻碍。因此,可以推测虽然华山松种群整体上表现为增长趋势,但是其自然更新仍然存在一定的隐患。这与裸果木[28]、羽叶丁香[9]和水曲柳[29]的种群特征类似。
静态生命表分析表明第Ⅰ、Ⅱ龄级个体生命期望寿命较高,但实际存活量较少,由此推断华山松种群幼龄个体受到了较强的外界干扰。实地调查中发现样地经常受到周围村民放牧、薪炭柴采集等活动的影响,幼苗损失现象极为严重。已有研究发现,森林更新过程中幼苗阶段是最重要和敏感的阶段[19],种群更新的限制因子主要是有限的空间和资源[30]。张小鹏等[31]研究表明,华山松幼苗储量大,但幼苗向幼树过渡阶段死亡率较高。这是由于华山松属于松属植物,是生态系统中的先锋树种,其生长发育过程需要良好的光照条件。虽然华山松种群幼苗丰富,但是随着幼苗的生长,林下相对郁闭的环境不能满足幼树对光照和营养空间的需求,导致了强烈的“自疏和他疏”现象。因此,可以推测由于华山松幼苗的竞争力弱以及环境的筛选作用,导致幼龄林个体无法进入下一龄级,这已经成为华山松种群更新和发展的“瓶颈”。
3.2 华山松种群的动态变化趋势
种群静态生命表和存活曲线可以直观地描述种群的现实状况,揭示种群与环境之间的适应机制[32]。静态生命表表明,华山松种群的死亡率和消失率变化趋势基本一致,前期缓慢增加,中期加速增长,末期急速上升。华山松种群的存活曲线趋近于Deevey-Ⅱ型,在第Ⅷ龄级之前死亡率相对平稳,之后死亡率突然上升,说明华山松种群进入了生理衰亡期,老龄林个体对空间和营养资源的竞争力下降,植株个体逐渐衰老。生存函数是任意时刻的函数,比存活曲线能更能直观和具体地反映种群结构和动态变化[33]。
生存分析表明,随着龄级的增加,华山松种群生存率逐渐降低,累计死亡率逐渐升高,两者呈相反的变化趋势;死亡密度曲线变化平缓,且稳中有降,而危险率曲线则呈逐渐增加的趋势,总的来说,危险率高于死亡率。这是由于老龄华山松个体数量较少,尽管死亡率很高,但是死亡密度呈降低趋势。综合4个生存函数分析,华山松种群整体表现为前期平稳、中期增长、后期衰退的动态特征,这可能是由于幼龄林阶段华山松的生物学特性和其生境共同导致了高死亡率,仅有小部分个体能够进入下一龄级;随着个体的发育,对生存空间、营养物质和光照等环境条件的需求逐渐增加,种间和种内的竞争加剧,导致生长脆弱的个体不能满足自身所需的资源而被淘汰,死亡率较高;老龄林阶段,由于个体进入了生理衰退期,对外界干扰的抵抗力变弱,种群迅速走向消亡。
时间序列分析表明,在未来2、4、6、8龄级后,Ⅰ~Ⅲ龄级种群个体数呈逐渐减少的趋势,Ⅴ~Ⅸ龄级种群个体数则呈逐渐增加的趋势,这说明种群在短期内不会出现毁灭性的衰退,仍能保持增长趋势,但是由于幼苗、幼树数量不足,不能充分补充种群中、壮龄阶段个体数量,最终必然会导致种群出现龄级断层,逐渐衰退。
3.3 种群更新及管护建议
植物种群更新是种子生理特征、幼苗更新能力与环境相互作用的过程[18],受种源、传播和建成的影响[34]。张小鹏等[31]研究发现,影响华山松幼苗定居的关键因子是土壤含水量和凋落物厚度,决定幼苗向幼树成功过渡的关键因子是光照。研究区内华山松种子丰富、土壤含水量高和凋落物层薄保证了幼苗补给充足,但是由于研究区周边农民的人为干扰,幼苗经常受到牲畜的破坏;同时,研究区内华山松密度较高,林分郁闭度度高,林内光照不足,影响了幼苗向幼树的过渡。由此可见,华山松自身的生物学特性、生境条件和人为因子影响着华山松的更新。
综上分析,仅依靠华山松林下天然更新难以维持其种群稳定性,加强幼苗和幼树的保护是保持华山松种群增长的关键。因此,在森林经营过程中需清除林下活地被物、凋落物等,改善幼苗的生存环境,提高幼苗的定居成功率;改善林内光照条件,建立小面积林窗,为更新层个体的发育创造有利环境条件,提高幼苗、幼树的成活率;加强管理,防止周围村民采伐和过度放牧等人为干扰行为对幼苗的破坏,为华山松种群的持续稳定发展提供保障。
