神经肽Y 系统特征及其鱼类生理调控研究进展
2020-06-19项骁炜王天明杨静文
梁 晶,张 炳,项骁炜,王天明,杨静文
(浙江海洋大学海洋科学与技术学院,国家海洋设施养殖工程与技术研究中心,浙江舟山 316022)
神经肽Y(neuropeptide Y,NPY),是1982 年由TATEMOTO,et al[1]等人首次从猪脑组织中提取得到的一种由36 个氨基酸组成的小分子单链多肽,分子量为4.271 kDa。随后其又在鸟类、两栖动物和鱼类中被分离出来。NPY 在结构上与多肽YY(peptide YY,PYY)、胰多肽PP(pancreatic polypeptide,PP)、胰多肽Y(pancreatic peptide Y,PY)相似,故同属于胰多肽家族[2]。鱼类主要分泌NPY 和PYY,只有部分硬骨鱼能够分泌PY[3],包括罗非鱼Oreochromis mossambicus[3]、黑鲈Micropterus salmoides[4],而PP 目前发现只存在于四足动物[5]。
在脊椎动物中,NPY 广泛分布在中枢神经系统和外周神经系统,其通过激活靶组织细胞表面的NPY受体(neuropeptide Y receptor,NPYR),介导下游信号通路,进而促进动物摄食和生长,亦在调节体温、能量代谢、生殖发育、心血管收缩以及压力响应与酒精代谢等方面发挥作用。目前在硬骨鱼类的研究中发现,NPY 系统可以促进摄食,也能刺激生长激素(growth hormone,GH)和胃促生长素(ghrelin)的释放,同时还能调节促性腺激素(gonadotropic hormone,GtH)和促黄体激素(luteinizing hormone,LH)的释放,被视作调节摄食、生长和生殖三者关系的重要协调因子。
1 神经肽Y 的结构及分布
1.1 神经肽Y 的结构
NPY 成熟肽的一级结构十分保守,其N 端有一个酪氨酸残基,C 端有一个酪胺酰胺残基,C 端的酰胺化对于NPY 的生物活性具有重要作用[6]。N 端的酪氨酸残基既能帮助稳定NPY 的三级结构又与NPY 及其受体的结合有很大关联[7],C 末端区域具有发夹样(hairpin)的空间结构,一般被称为PP 折叠(PP-fold)[2],此结构是由脯氨酸延伸构成的脯氨酸螺旋(prolinehelix)通过一个转折(turn)后与2 个酪氨酸构成的α 螺旋交叉形成,而α 螺旋结构的稳定性取决于pH 的变化。
从脊椎动物的NPY 核心序列比对可以看出,不同物种NPY 的氨基酸序列均有高度同源性[8]。如图1所示,在不同物种中,非洲爪蟾Xenopus laevis与加州鲈Micropterus salmoides的NPY 成熟肽序列最多仅存在6 个不同的氨基酸(图1)。基于胰多肽家族中NPY、PY、PYY 成熟肽氨基酸序列,构建了16 个不同物种的NJ 系统进化树。进化树结果表明,脊椎动物中胰多肽家族的NPY 氨基酸序列同源性与其分类地位基本吻合(图2)。
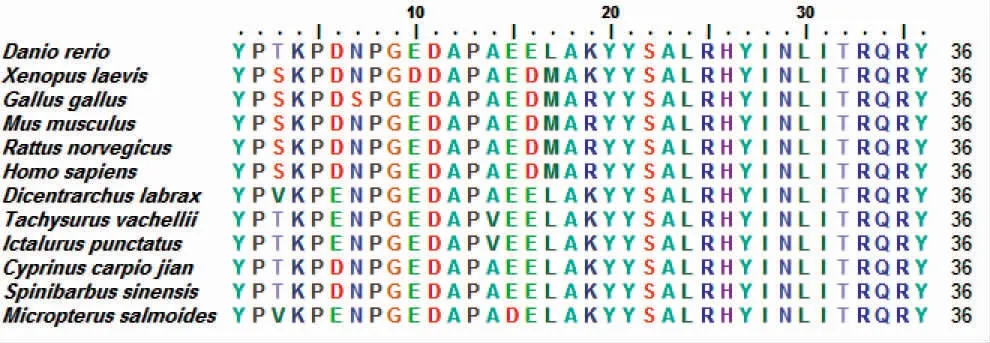
图1 不同物种的NPY 成熟肽序列比对[9]Fig.1 NPY mature peptide sequence alignment of different species

图2 不同物种中胰多肽家族成熟肽氨基酸序列的系统发育树Fig.2 Phylogenetic trees of aminoacids sequences of pancreatic peptide family mature peptides in different species
1.2 神经肽Y 的分布
NPY 作为一种神经递质参与体内多种生理功能的调节,在生物体中分布十分广泛。NPY 主要分布在神经系统中,在中枢神经系统方面,NPY 在端脑、视前区、视顶盖以及丘脑中含量十分丰富,尤其在下丘脑弓状核中常能发现较高浓度的NPY,调控下游因子及激素的合成与释放,从而促进生物体的摄食行为[10];另外,NPY 在海马组织内含量也非常丰富,大部分产生于γ-氨基丁酸中间神经元,主要起到降低神经元兴奋性的作用[11]。而在外周神经系统方面,NPY 主要分布在交感节后神经元以及肾上腺髓质等,并且NPY 常与交感神经递质去甲肾上腺素共存[12]。
2 神经肽Y 受体
NPY 在不同生物和器官中可执行不同的生理功能,主要通过激活其不同的受体(NPYR)而得以实现。NPYR 作为NPY 在细胞膜上的靶向受体,属于G 蛋白偶联受体(G protein-coupled receptors,GPCR)家族,具有保守的7 次跨膜螺旋结构,是生物体内最大的细胞膜受体蛋白家族,其N 端在膜外主要负责与信号递质结合,C 端在胞内主要介导各类激酶的活化,在将细胞外信号传递到细胞内效应分子的过程中发挥了重要作用[1]。目前研究发现神经肽Y 受体共有8 种亚型受体(Y1~Y8),早期脊椎动物的基因组在进化中,由一个NPY 受体基因通过2 次局部复制产生了Y1、Y2 和Y5 3 个受体亚家族,包括Y1 亚族受体(Y1、Y4、Y6、Y8)、Y2 亚族受体(Y2、Y7)和Y5 亚族受体(Y5)[13-15]。在进化过程中,哺乳动物已经丢失了Y7 与Y8 2 种受体,目前只克隆获得Y1、Y2、Y4、Y5 和Y6 5 种亚族受体[16];在硬骨鱼中,一直未发现Y5 和Y6 2种亚族受体,猜测二者可能均已丢失,另外,Y1 受体仅在斑马鱼中被发现,Y3 受体目前只是基于生物信息学的预测,还未在任何物种中克隆出来[11,17](图3)。不同基因座上的独立基因编码不同亚型的NPYR,因此NPY 在分布、表达、生理功能以及与受体之间的结合能力上差异明显,也是该系统研究的重要内容[18]。

图3 不同物种中NPY 受体氨基酸序列的系统发育树Fig.3 Phylogenetic tree of NPY receptor amino acid sequences in different species
3 NPY/NPYR 系统介导的生理功能
NPY 经过与不同的亚型受体结合,激活信号转导通路,触发多种生理功能。NPYR 被激活后主要活化对PTX(百日咳毒素)敏感的Gi蛋白和对PTX 不敏感的Gq蛋白。在多种情况下,NPYR 介导的信号能抑制腺苷酸环化酶(AC)活性以降低3’,5’一环腺苷酸(cAMP)的合成;NPY 能通过依赖或不依赖于肌醇三磷酸路径活化胞内钙库增加钙离子水平;NPYR 还能通过与膜上的Gq蛋白结合活化磷脂酶C(PLC),刺激二酰甘油(DAG)与三磷酸肌醇(IP3)生成,调节钙离子通道和钾离子通道[19],从而实现对生物体生理功能的调控。目前研究认为:NPY 主要通过Y1,Y2,Y5 受体发挥促食欲作用[20-22]。Y2 还与记忆功能相关[23],Y4 可能与食欲减退功能有关[24],而目前研究还没有证实Y6 受体的功能[25-26]。
3.1 NPY/NPYR 对鱼类摄食行为的调节
鱼类摄食行为的调节是一个非常复杂的过程。摄食行为是由中枢神经系统以及外周神经系统相互协调并在下丘脑整合进行调节的。NPY 作为下丘脑食欲调节网络中一种十分重要的内源性促食欲因子,对生物体能量的摄入、消耗、储藏及平衡起到至关重要的作用。在下丘脑的摄食调节中,弓状核是脑肠肽(ghrelin)在中枢神经起作用的主要位点。Ghrelin 的神经元与下丘脑部的NPY 或刺鼠相关肽(AgRP)以及阿黑皮素(POMC)的神经元胞体以及突触相连接,从而实现对生物体的摄食调节(图4)[27]。有研究对金鱼Carassius auratus 进行饥饿处理,发现饥饿24 h 后,金鱼下丘脑组织中NPY mRNA 表达显著上升,饥饿72 h后在金鱼各脑区均有NPY mRNA 的表达,尤其是大脑端脑的视前区及丘脑顶盖部分发现表达大量NPY mRNA,在饥饿后重新喂养金鱼,NPY mRNA 的表达很快恢复正常[28-29],由此表明,NPY 在金鱼的摄食调控中发挥了重要作用。并且此结果在斑马鱼Danio rerio[30]、鲢鱼Hypophthalmichthys molitrix[31]、银大麻哈鱼Oncorhynchus keta[32]、南方鲇Silurus meridionalis[8]以及鲑鱼Oncorhynchus kisutch[33]中得到证实,说明NPY 是参与鱼类摄食调节中重要的神经内分泌信号。
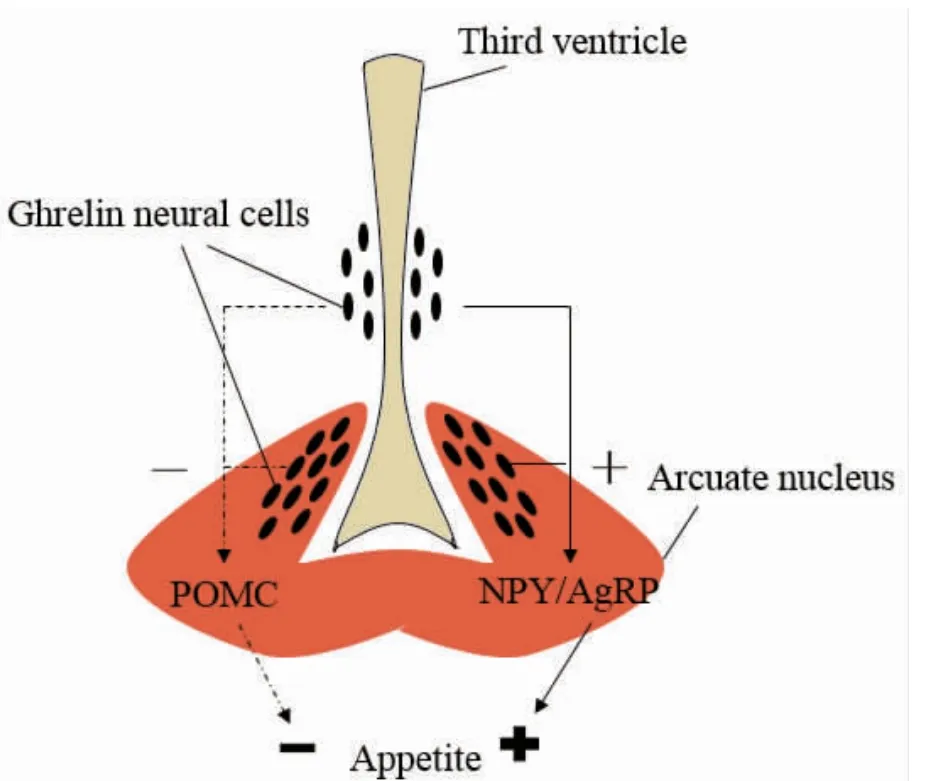
图4 下丘脑部的摄食调节神经网络[26]Fig.4 Network of neural feeding regulation in hypothalamus
由于NPY 必须与相应NPYR 结合才能发挥相应的生理功能调节作用,因此NPYR 对鱼类摄食行为的调节与其特异性受体密切相关。研究人员在金鱼脑室注射NPY 的Y1,Y2 及Y5 受体激动剂2 h 后,发现Y1 和Y5 2 种受体可增加金鱼的摄食能力;而Y2 受体对金鱼的摄食能力没有作用[34]。还有研究人员对虹蹲Oncorhynchus mykiss脑室注射NPY 的Y1,Y2 及Y5 受体激动剂后,发现Y1 及Y2 受体能够促进虹鳟的摄食活动;但注射Y5 受体激动剂的虹蹲完全不受影响[35]。在斑马鱼中,通过在脑室中注射NPY 的Y1 受体拮抗剂,则完全抑制了NPY 的作用[30]。推测不同鱼类中参与摄食调节的NPYR的种类也有不同,比如金鱼的摄食活动是由Y1 和Y5受体介导的,而虹鳟的摄食活动是由Y1 及Y2 受体介导的。
3.2 NPY/NPYR 对神经内分泌的调节
NPY 分布于下丘脑弓状核、室旁核、室周核、正中隆起等多个区域,这些区域又是许多内分泌细胞存在的部位,它们相互联系共同调节机体复杂的生理活动。已有的研究证明,NPY 对生长激素(GH)、促性腺激素释放激素(gonadotropin releasing hormone,GnRH)、催乳素(prolactin,PRL)、促甲状腺激素(thyroid stimulating hormone,TSH)、黄体生成素(luteinizing hormone,LH)、生长抑素(somatostatin,SS)、促肾上腺皮质激素释放激素(corticotropin releasing hormone,CRH)和促性腺激素(GtH)等内分泌激素都有一定的调节作用[35-37]。
3.2.1 NPY/NPYR 对下丘脑-垂体-性腺轴(hypothalamic-pituitary-gonadal axis,HPG 轴)调节的影响
研究表明下丘脑-垂体-性腺轴释放的激素分别为:GnRH、LH/卵泡刺激素(follicle-stimulating hormone,FSH)和性激素。HPG 轴的主要功能是调控维持性的发育和性别特征。GnRH/GnRHR 信号系统是生殖发育HPG 调控轴的关键组成。早在1988 年,在研究NPY、GnRH 在兔子以及小鼠中表达时就有发现,切除卵巢的兔子及小鼠,NPY 不能刺激GnRH、LH 的表达,而不切除卵巢,则能够刺激GnRH、LH 的表达,结果表明NPY 对于GnRH、LH 的调节作用必须依赖于卵巢的存在[38-39]。还有研究发现,在哺乳动物及鱼类脑室注射NPY 对GH、TSH 和LH 的释放都有一定的影响[40]。在鱼类中,研究者借助于免疫化学分析等研究方法,在虹鳟、褐鳟Salmo trutta、以及蟾胡鲶clarias batrachus中均发现NPY 与GnRH 的调节有紧密的联系[41-42]。利用双重免疫标定法,研究人员发现香鱼Plecoglossus altivelis的NPY 免疫阳性神经纤维与视前区的GnRH 分泌细胞的位置接近且并列一起[43]。GAIKWAD,et al[42]研究蟾胡鲶发现前脑NPY 从嗅觉上皮细胞扩展至前脑垂体等不同的神经轴调节GnRH 的分泌,推测NPY 能够影响鱼类GnRH 的表达。目前研究认为NPY 通过激活Y1 受体来促进GnRH 的分泌,尤其是直接增强GnRH 的分泌从而促进LH 从促性腺激素释放激素靶细胞的释放,在性激素正常或较高情况下,NPY 使下丘脑GnRH 的分泌增多,而在较低情况下,则抑制其分泌,且NPY 对于GnRH 抑制主要是通过Y5 受体来介导的[44]。
3.2.2 NPY/NPYR 对下丘脑-垂体-肾上腺皮质轴(hypothalamic-pituitary-adrenal axis,HPA 轴)调节的影响
NPY 分布于下丘脑的室旁核神经元纤维中,而室旁核是HPA 轴活动的直接控制部位,HPA 轴是与应激反应关系最为密切的内分泌轴。有研究对大鼠进行水上漂浮以及高强度运动等实验,来证实大鼠在受到应急刺激时,室旁核的NPY 神经元纤维可支配下丘脑分泌2 种重要的促肾上腺皮质激素释放激素(CRH)和精氨酸加压素(AVP),并刺激垂体分泌促肾上腺皮质激素(ACTH),最终使肾上腺皮质激素分泌增多[45]。由此可知NPY 对HPA 轴的活动有着一定的调控作用。另外HPA 轴也可影响NPY 系统,如CRH 通过负反馈抑制NPY 的表达[8]。皮质醇作为一种应激激素,能使生物体对压力做出有效反应。在鱼类中,有研究人员在金鱼腹腔内注射皮质醇,72 h 后发现端脑视前区NPY 分泌增多,CRF mRNA 水平的降低。结果表明,前脑NPY 和CRF 可能在HPA 轴的调控作用中发挥作用[46]。
3.3 NPY 的其他作用
除上述NPY 对鱼类摄食行为、HPG 轴、HPA 轴调节外,NPY 对心血管系统、呼吸系统、循环系统以及消化系统也有一定的作用。NPY 可以与Y1 亚型受体结合使血管收缩,血压升高,或者作用在中枢升高外周阻力等,很少量的NPY 就可引起缓慢且持久的血管收缩[47]。有研究人员在鳗鲡Anguilla japonica[48]、雀鳝Atractosteus spatula[49]的胰腺中发现NPY 免疫阳性细胞;在大西洋鳕鱼Gadus morhua[50]及猫鲨Chiloscyllium punctatum[51]的消化管道的神经纤维以及内分泌细胞中有NPY 免疫反应,推测NPY 可能参与鱼类胃肠道的消化和吸收调节、胰腺的分泌活动调节以及与心血管活动调节。
4 神经肽F 的生理功能
神经肽F(NPF)发现于无脊椎动物,是脊椎动物中NPY 的同源多肽,NPF 也主要分布在中枢神经系统,另外其在肝脏、食管、性腺等器官中也有部分表达,对生物体生理调节机制起到重要作用[52]。无脊椎动物中最早在莫尼茨绦虫Monieza expansa[53]中发现与脊椎动物NPY 同源的序列,因其序列螺旋C 末端的酪氨酸Y 被苯丙氨酸F 替换(RPRFamide),所以将NPY 的同源肽称为NPF(neuropeptide F,NPF)。NPF 有很多与NPY 类似的调控功能,包括摄食[54]、学习与记忆[55]、繁殖[56]、压力反馈[57]等。目前已经在加州海兔Aplysia californica[58]、静水椎实螺Lymnaea stagnalis[59]、枪乌贼Loligo vulgaris[60]、帽贝Patellace[61]、太平洋牡蛎Crassostrea gigas[62]等物种中获得了NPF 的全长序列。自1992 年RAJPARA,et al[58]在加州海兔中枢神经系统、食道神经细胞以及腹部神经细胞都有发现NPF 的表达后,JING,et al[63]在加州海兔颊和脑神经节中发现,在给加州海兔注射NPF 后可使其产生饱食感,进而减少进食量,说明海兔NPF 与哺乳动物中NPY 的促进摄食作用正好相反。研究人员在给静水椎实螺注射NPF 后发现,NPF 对其摄食行为没有影响,却能够抑制其生长与繁殖,这说明NPF 可能不参与静水椎实螺的摄食调控而参与其生长繁殖调控过程[59]。
5 神经肽Y 及其受体的研究前景展望
NPY 在生物体内广泛存在,在中枢以及外周都起着非常重要的调节作用。近年来随着NPY 及其受体研究的不断深入,其作用机理及参与的生理调控网络被进一步的阐释。目前研究鱼类中NPY 对生物体的摄食、生长繁殖以及神经内分泌的调节等生理机制已有了初步的了解,但尚不够清晰。例如,1、在鱼类中,NPY 为何具有如此广泛的生理调控功能?2、NPY 为何能够在不断地进化中保持其稳定性?3、Y7 与Y8 两种亚型受体仅在鱼类及两栖动物中存在的意义?4、在鱼类幼年期与成年期NPY 的作用是否改变?等这些科学问题都有待更多的研究结果给予更充分的支持和说明。对鱼类中NPY 的进一步深入研究,将有助于我们深入了解NPY 在鱼类的生长、发育、生殖等多种生理调控相关的神经内分泌系统中的作用,也将为系统地认识NPY 系统在不同动物进化演变中的功能角色转变提供重要参考。
