水稻稻瘟病抗性基因Pik-2的遗传变异及进化分析
2020-06-12牛玉芳肖应辉
牛玉芳, 肖 贵, 吴 俊, 柏 斌, 周 波,3, 肖应辉
(1. 湖南农业大学 农学院, 长沙 410128; 2. 杂交水稻国家重点实验室 湖南省杂交水稻研究中心, 长沙 410125; 3. 国际水稻研究所, 菲律宾 马尼拉 0900)
由子囊真菌(Magnaportheoryzae)引起的稻瘟病,是在世界范围内广泛发生的水稻病害之一。在对抗稻瘟菌不断变异的进化过程中,水稻形成了复杂多变的抗病机制和丰富多样的抗性基因。已有文献概述了编码NBS-LRR蛋白的Pb1、Pia、Pib、Pid3、Pik、Pik-h/Pi54、Pik-m、Pik-p、Pish、Pit、Pita、Piz-t、Pi1、Pi2、Pi5、Pi9、Pi25、Pi36、Pi37、Pi56、Pi63、PiCO39、Pi50、Pi65(t)和Pigm,以及分别编码B-Lectin蛋白激酶的Pid2和富含脯氨酸蛋白的pi21等28个稻瘟病抗性基因[1]。最近又报道了不同于以往抗性机制的新基因Ptr[2]、Bsr-d1[3]和bsr-k1[4]。
位于水稻第11染色体的稻瘟病抗性基因座Pik,是水稻稻瘟病抗性基因成簇分布的3个热点区域之一。Kiyosawa[5]最先报道了Pik、Pikh、Pikm、Pikp和Piks5个等位基因,Pan等[6]在该位点鉴定到抗性反应不同于前5者的Pikg,后来陆续鉴定出Pi1[7]、Pi7[8]、Pilm2[9]、Pi43[10]、Pi46[11]和Pi47[1]等多个新基因。Ashikawa等首次报道Pik抗性由Pikm-1、Pikm-2两个NBS-LRR类基因共同控制,Pik功能的实现需要两个NBS-LRR蛋白共同作用,相应的无毒蛋白AvrPik只与其中的Pik-1蛋白直接互作[12],Pik-2与Pik-1能够形成异源二聚体[13]。
随着Pik座位中基因的不断克隆,其序列差异与进化研究也取得显著进展。Ashikawa等[14]克隆了Kanto51中的稻瘟病抗性基因Pik-KA,与大多Pik等位基因相比,该基因的Pik-1中编码决定Pik的抗性特异性的CC结构域序列发生了显著变异,而其Pik-2与其他Pik基因编码的氨基酸序列完全一致,暗示Pik基因中的Pik-2非常保守。Constanzo等用特异性PCR引物扩增测序分析了15个美国水稻品种Pik-m基因的DNA序列,发现核酸多态性都发生在Pikm-1的DNA序列,而Pikm-2等位基因即使在亲缘关系甚远的品种中仍然保守[15]。目前关于Pik-2基因的多样性及其抗性功能方面研究鲜有报道。
为了深入研究Pik-2的遗传变异机制,本研究对来自不同国家的400份非洲稻和400份亚洲稻在Pik-2基因编码区进行了测序分析,拟通过分析Pik-2核苷酸水平上的多态性,明确非洲稻种和亚洲稻种Pik-2序列变异特点及其与地理分布的关系,为进一步挖掘非洲和亚洲水稻品种中蕴含的稻瘟病抗性基因提供参考。
1 材料和方法
1.1 供试材料
400份非洲稻(Oryzaglaberrima)和400份亚洲稻(Oryzasativa)种质取自国际水稻所种质资源库。水稻种质CO39、丽江新团黑谷(LTH)、日本晴、Taipei309、IR64、IRBLKp-K60和IRBLK-Ka由杂交水稻国家重点实验室提供,其中IRBLKp-K60和IRBLK-Ka为含有Pikp和Pik-KA基因的单基因系。
1.2 水稻基因组DNA的提取及检测
取3~4叶期水稻植株的新鲜组织100 mg,装入2 mL离心管中,用液氮研磨成粉末。采用CTAB法进行DNA基因组的提取[16]。
1.3 Pik-2同源基因的PCR扩增与测序
分析已克隆的9个Pik等位基因的Pik-2 LRR区域,发现氨基酸多态性存在于该基因编码蛋白的627与754位点[5,14,17-19],相应的DNA碱基序列位于1879~2457 bp之间,因此根据已知的Pik-2基因的保守序列设计引物对(RGA5-F:5′-ACAAAAGGCCTGAACTTGGCTCAAGTG-3′;RGA5-R: 5′-TCATGCAGTGACGATGCCATCAAC-3′),由北京擎科新业生物技术有限公司合成。
PCR反应体系包括:0.25 μLTaqDNA 聚合酶(5 U/μL),5.0 μL 10×bufffer,4.0 μL dNTPs(10 μmol/L),1.0 μL 正向和反向引物(10 μmol/L),2 μL 模板 DNA,加 ddH2O至50 μL。PCR反应程序为:94 ℃预变性5 min;98 ℃变性20 s,58 ℃退火30 s,72 ℃延伸2 min,35个循环;72 ℃再延伸10 min。PCR扩增产物用1%的琼脂糖凝胶电泳,电泳后于紫外投射仪下检测条带合格后,PCR产物委托北京擎科新业生物技术有限公司测序。
1.4 序列分析
用Sequencher 5.4.6软件进行序列比对分析。用软件DNAMAN、BioEdit以及DNASP进行单倍型以及差异性分析。然后用软件MEGA 6.06构建系统进化树。变异频率以某位点碱基变异的种质份数/总种质份数×100%表示。
2 结果与分析
2.1 Pik-2序列扩增及变异分析
采用特异性引物RGA5扩增Pik-2的LRR区域,在含有Pik-2的水稻品种中均能扩增出特异性单一条带(图1),而不含Pik-2序列的水稻品种中均未出现任何条带,表明该引物对能有效扩增特异性的Pik-2序列。利用该引物扩增400份非洲稻和400份亚洲稻的全基因组DNA,分别获得336份非洲稻和306份亚洲稻的Pik-2序列。
对Pik-2基因碱基位置1879~2457的核苷酸变异分析发现,在非洲稻中共检测到113个变异位点,包含20个转换与94个颠换位点,这些位点的变异频率为0.30%~99.70%,平均突变频率为43.11%。在亚洲稻中共检测到78个变异位点,包含18个转换与65个颠换位点,变异频率为0.33%~99.35%,平均突变频率为37.68%。值得一提的是,在非洲稻和亚洲稻中突变频率最高的位点均发生在1879碱基位点,其突变频率分别为99.70%和99.35%。数据显示,与亚洲稻对比,Pik-2基因在非洲稻中的变异更丰富。

图1 特异性引物鉴定
2.2 Pik-2单倍型和遗传多样性分析
基于水稻稻瘟病抗性基因Pik-2 LRR结构域部分序列的核苷酸差异,将336份非洲稻划分为17种单倍型(图2),分别命名为OG-H1—OG-H17;将306份亚洲稻划分为12种单倍型,命名为OS-H1—OS-H12。
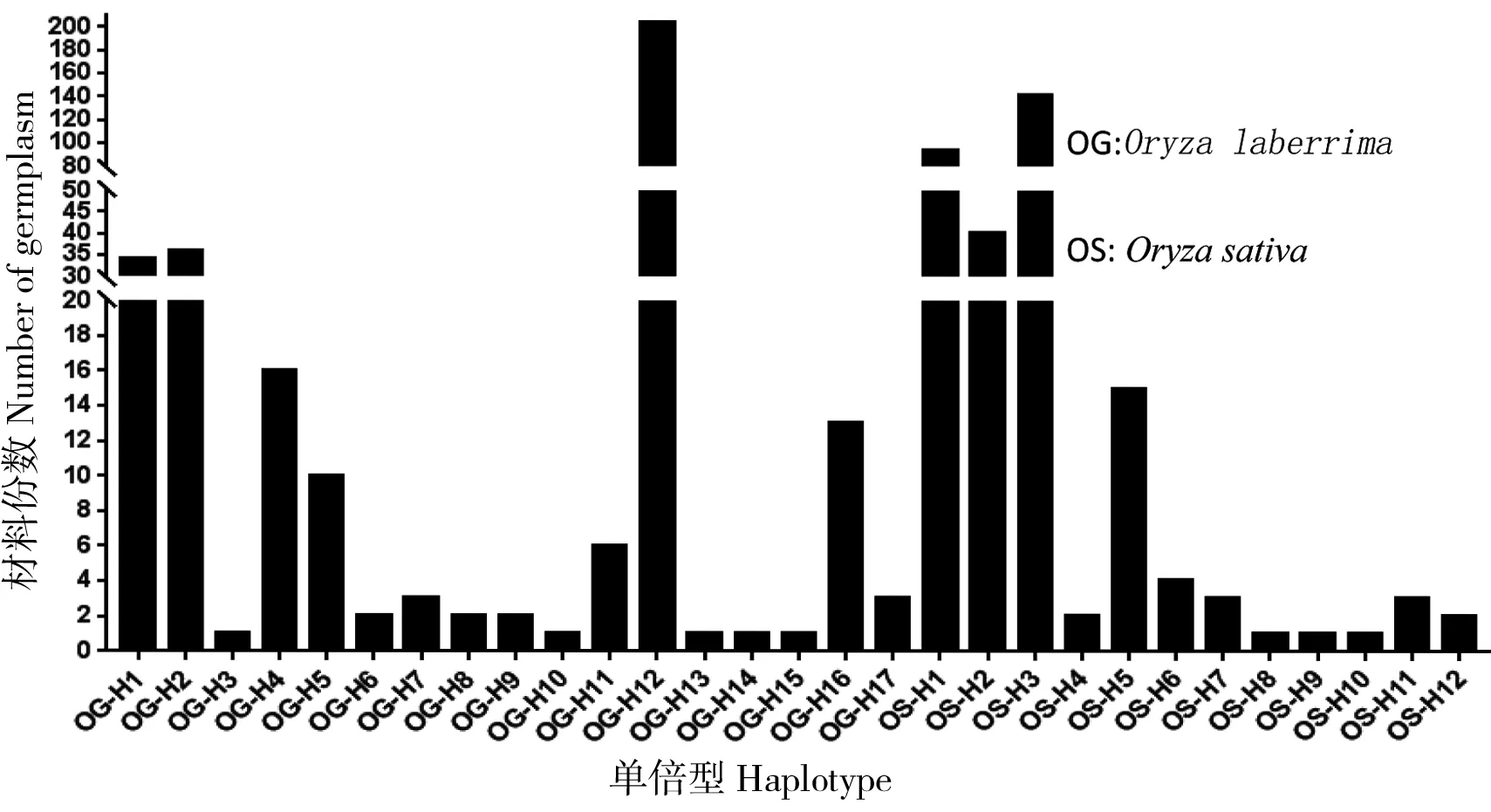
图2 非洲稻和亚洲稻的单倍型分布
非洲稻中,单倍型OG-H12为优势单倍型,有204份水稻种质属于该单倍型。OG-H12与OG-H13仅有2个碱基位点(一个氨基酸位点)差异,而OG-H13却只存在1份非洲稻中。亚洲稻中,单倍型OS-H1和OS-H3为优势单倍型,分别在93和141份材料中检测到这两个单倍型(表1)。
17种非洲稻单倍型共包含51个氨基酸位点的变异,其中单倍型OG-H2、OG-H4、OG-H5和OG-H6中碱基变异并未引起氨基酸变化,又如单倍型OG-H1与Pik-2相比有14个碱基变异位点,而仅在697、709、718、737和800 5个位点的氨基酸发生改变(表2),说明非洲稻不同单倍型的碱基变异大多为同义突变。12种亚洲稻单倍型的78个碱基位点改变却导致41个氨基酸位点变异。非洲稻和亚洲稻没有相同的单倍型,但有36个相同的氨基酸变异位点。结果显示,Pik-2的LRR区域在非洲稻与亚洲稻中可能存在不同的进化模式,非洲栽培稻的变异更为丰富但同义突变较多,亚洲栽培稻的变异较少但多为非同义突变。
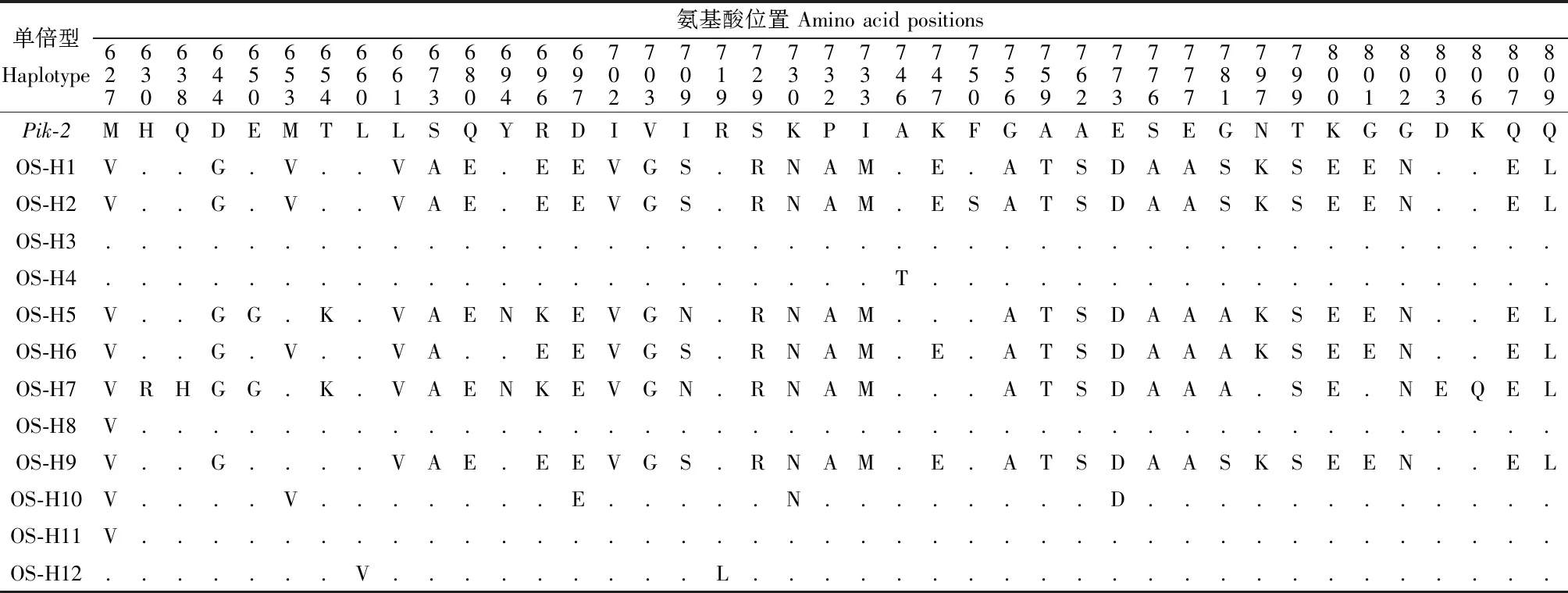
表1 亚洲稻 Pik-2不同单倍型氨基酸序列变化
2.3 Pik-2单倍型在非洲与亚洲各地区分布
非洲稻Pik-2基因的17种单倍型在西非国家都有分布,中非国家只有OG-H1、OG-H2与OG-H12 3种单倍型,南非分布的单倍型有OG-H5、OG-H11与OG-H12(图3-A),总体上呈现出大聚集、小分散的分布特点。非洲水稻种植主要集中在西非国家,本研究收集的非洲稻有95%来自西非国家,可能是导致上述结果的原因。在这些单倍型中,OG-H12分布最广泛,在大多西非国家中都存在该单倍型,部分单倍型只在某些特定国家中发现。如在非洲国家布基纳法索发现了3种特有的单倍型OG-H3、OG-H10和OG-H13,在尼日利亚发现2种特有的单倍型OG-H14和OG-H15,且这些单倍型都只存在其中一份水稻种质中。同时还发现只在特定国家多份水稻种质中存在的单倍型,如包含单倍型OG-H4的16份水稻种质均来源于尼日利亚,而3份OG-H17单倍型的水稻种质均来源于利比里亚。从非洲国家的单倍型分布来看,尼日利亚、马里和布基纳法索拥有较多类型的单倍型。
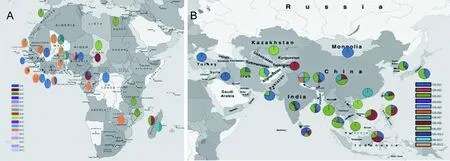
图3 非洲稻(A)和亚洲稻(B)不同单倍型的地理分布
注:图中圆环中不同扇形颜色代表不同的单倍型;扇形面积大小代表该单倍型在该地区的分布比例; 周边有黑色线条的圆环代表亚洲稻材料,无黑色线条者代表非洲稻材料
本研究中的亚洲稻来自63个国家或地区(图3-B),其中亚洲、非洲、美洲、欧洲和大洋洲的比例分别为70.3%、7.26%、15.84%、5.94%和0.66%。亚洲稻的12种Pik-2单倍型中,OS-H4、OS-H7和OS-H9 只分布于南亚国家,OS-H1和OS-H3在四大洲都有分布。亚洲稻的4个单倍型OS-H8、OS-H9、OS-H10和OS-H11都只在单份水稻种质中检测到,这些材料分别来自多米尼加、孟加拉、中国和不丹。与非洲稻相比,亚洲稻单倍型分布相对广泛。
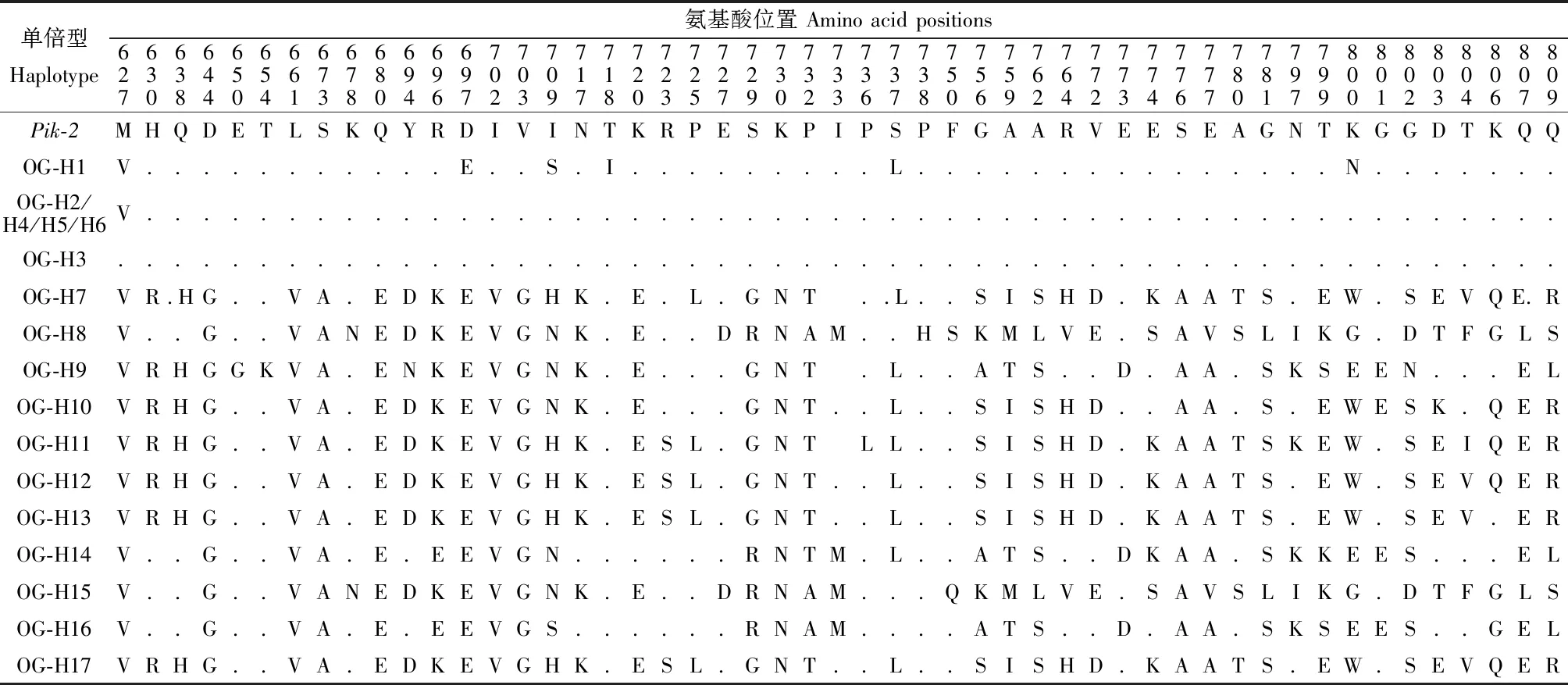
表2 非洲稻Pik-2不同单倍型氨基酸序列变化
2.4 Pik-2不同单倍型的进化分析
系统进化分析表明,取遗传距离为0.015为阈值,可以将非洲稻和亚洲稻的所有单倍型以及已经克隆的Pik-2 等位基因(Piks-2、Pikm-TS-2、Pik-KA-2、Pik-KU-2、Pik-LTH-2、Pikp-2、Pikh-2、Pi7-2和Pi1-6)分为4类(图4)。其中,Pik-2等位基因与非洲稻6个单倍型(OG-H1、OG-H2、 OG-H3、OG-H4、OG-H5、OG-H6)及亚洲稻6个单倍型(OS-H3、OS-H4、OS-H8、OS-H10、OS-H11、OS-H12)归为一类,这一类亚洲稻和非洲稻种质与已报道的Pik-2基因往往只有1~2个编码氨基酸的差异。第2类包括的单倍型有OG-H12、OG-H17、OG-H7、OG-H13、OG-H11和OG-H10,这一类群均为非洲稻种质,627-703、717-720、727-733、737-773和799-803位置的氨基酸序列完全一致。第三类包括的单倍型有OG-H8和OG-H15,这两个单倍型在碱基2247-2249和2251-2253处都发生了碱基缺失变异。第4类包括的单倍型有OS-H5、OG-H9、OS-H7、OG-H14、OG-H16、OS-H9、OS-H1、OS-H2和OS-H6,在627、660、661、697、703、730、756、759、762、773、776、777、800、807和809号位置的氨基酸完全一致。
进化树分析表明,依据水稻稻瘟病抗性基因Pik-2 LRR结构域核苷酸单倍型分类,不能将非洲稻与亚洲稻明确分组,暗示非洲稻与亚洲稻在Pik-2 LRR结构域的进化可能相互独立,在各自进化的过程中又形成了大致相似的单倍型。如第一类中的亚洲稻单倍型OS-H3和非洲稻单倍型OG-H5、OG-H2的相似系数极高,只有627、630和638处3个氨基酸的差异。

图4 Pik-2单倍型进化树分析
3 讨论与结论
目前已有30多个稻瘟病抗性基因被克隆,进一步挖掘新的抗性基因难度越来越大,不断挖掘已知抗性基因的不同等位基因,用来对抗不同生态区域复杂易变的稻瘟病生理小种,将是今后稻瘟病抗性基因资源挖掘的主要方向。已报道克隆的稻瘟病抗性基因大多成簇分布,多基因簇的序列高度相似。例如,Pi2、Pi9以及Piz-t互为复等位基因,对不同国家的稻瘟病菌株表现出广谱抗性;3个基因的序列高度一致,Pi2和Piz-t只有8个氨基酸差异,Pi2、Piz-t与Pi9分别有43和46个氨基酸差异[20-21]。许多保守结构在新基因或者等位基因挖掘中具有重要作用,利用已知基因的保守序列,设计特异性功能分子标记,结合PCR及遗传分析等多种方法即可克隆目标基因的等位基因。因此,对这些抗性基因的遗传多样性分析,有助于挖掘抗谱更广的新等位基因。
Pik基因座是稻瘟病抗性等位基因数目最多的位点[22],且这些等位基因对中国不同稻区的稻瘟病生理小种大多具有较广谱及持久的抗性[23]。目前多数研究认为Pik-2是Pik-1基因的“helper”,协助Pik-1基因行使抗病功能,对Pik-2功能的认识往往是基于Pik-2序列保守性而设想的,这是因为迄今克隆的9个Pik-2等位基因只在4个氨基酸位点有多态性[24]。本研究在336份非洲稻和306份亚洲稻中分别检测到Pik-2基因的113和78个碱基变异,相应51和41个氨基酸位点变异,表明Pik-2在非洲稻和亚洲稻种质资源中存在比预期更为丰富的遗传多样性,这一研究将有助于了解和利用不同种质资源中Pik位点丰富多变的基因型。
本研究根据Pik-2等位基因碱基变异的多样性,将非洲稻和亚洲稻分别划分为17和12种单倍型。有些稀有单倍型只在单独或者少量种质中发生变异,这些种质将是进一步挖掘Pik-2新等位基因的重点。非洲稻Oryzaglaberrima病虫害抗性和耐高温干旱能力较亚洲稻有明显优势[25],同时保留了更多的遗传多样性。因此,挖掘和利用非洲稻中的Pik及其他稻瘟病抗性等位基因,对于亚洲稻区培育抗病品种具有重要价值。
