桂林喀斯特生境红背山麻杆克隆繁殖特征
2020-06-11周俊妞梅军林马姜明张雅君王海苗
周俊妞,梅军林,马姜明*,张雅君,王海苗,菅 瑞
(1. 广西师范大学 可持续发展创新研究院,广西 桂林 541006;2. 珍稀濒危动植物生态与环境保护教育部重点实验室(广西师范大学),广西 桂林 541006;3. 广西师范大学 生命科学学院,广西 桂林 541006)
喀斯特是一种具有溶解能力的水溶液对碳酸盐岩等可溶岩类的溶蚀过程,以及由此产生的地貌等现象的总称[1]。中国喀斯特地貌(karst landform)面积为3.443×106km2,约占全国总领土面积的35.9%[2]。由于喀斯特地区自然地理地质条件恶劣,生态系统脆弱,岩石裸露率高,土层浅薄,成土速度极慢[3],在长期的喀斯特作用下形成地表和地下双层空间结构,使得降落的雨水常通过地表溶岩裂隙、落水洞、洼地等直接快速渗入地下空间[4],导致喀斯特地区储水能力低,地表水匮乏,可利用的水资源短缺[5],植被生长缓慢。加之人为干扰频繁,植被严重退化,植物多样性降低,生态系统抗干扰能力和自我恢复能力减弱[6],导致石漠化现象日渐加剧。
克隆生长,也称克隆繁殖,是指在自然条件下由一个植株通过营养生长产生多个在遗传上一致、在形态和生理上独立或具有潜在独立生存能力个体的过程[7]。克隆植物除了具有较强的适应环境和抗干扰能力外,还具有很强的水平扩展能力,能有效地利用有限环境资源,迅速开拓新的生境,扩大种群,形成单优势种的立地[8-9],在植物群落演替中起着重要作用。克隆植物存在的广泛性[10]、空间的拓展性[11]、利用资源的高效性[12]等,使克隆植物更容易适应恶劣环境,具有较强的生存竞争能力[13],在生态系统中具有重要的地位。目前对克隆植物的研究主要集中在草本植物,如结缕草Zoysiajaponica、空心莲子草Alternantheraphiloxeroides、活血丹Glechomalongituba等,对木本植物的克隆繁殖研究较少,主要有火炬树RhusTyphina、沙棘Hippophaerhamnoides、胡杨Populuseuphratica,且研究地区主要针对高山、草原、沙漠等干旱环境,对于喀斯特地区克隆植物研究的报道较少。近些年,人们在克隆植物的构型、可塑性和克隆整合等方面取得了实质性研究进展[14-16],但对整合作用机理、整合时分株的耗益、整合发生和停止的时间与原因以及整合作用在生产上的应用等方面研究较少。
作为一种克隆植物,红背山麻杆具有繁殖周期短、拓展性强、利用资源高效、生长快等优点。相关研究表明,红背山麻杆是低蒸腾高水分利用率的树种[17]。在喀斯特石漠化地区种植红背山麻杆可利用其繁殖周期短、生长快等优点帮助改善石漠化地区的土壤性质,涵养水源、减少水土流失,防治石漠化恶化。本文通过研究红背山麻杆前后端水平侧根直径、分株数与地径的关系,拟为进一步探讨红背山麻杆克隆繁殖机制提供科学依据。
1 材料与方法
1.1 研究区自然概况
研究地为桂林市七星区朝阳乡西南村的典型喀斯特石山,地理坐标为110°34′~110°35′E,25°27′~25°26′N,海拔150~166 m,坡向西南,坡度30°(表1),土壤以石灰土为主,地表覆盖有大量碎石,裸露率达30%以上。桂林地处广西东北部,属中亚热带季风气候,雨量充沛,气候温和,年平均气温18.9 ℃,年均日照总时数1 550 h,全年无霜期300 d,年均降雨量1 900 mm,年平均蒸发量1 490~1 905 mm[18]。样地因人为干扰程度和地形因子的复合影响而发生次生演替,主要优势种有紫薇Lagerstroemiaindica和黄荆Vitexnegundo等灌木,金丝草Pogonatherumcrinitum、野鸡尾Onychiumjaponicum、千里光Senecioscandens等草本植物以及龙须藤Bauhiniachampionii等藤本植物。

表1 研究地区样地基本情况
1.2 样地设置及样本划分
研究地划分出6个红背山麻杆群落样地(10 m×10 m),其物种组成分属12科25属26种,群落中植物个体数为9 395株,其中红背山麻杆个体数为8 081株,占总个体数的86%,为绝对优势种。选取砍伐干扰克隆繁殖和无干扰克隆繁殖类型的红背山麻杆为研究对象,参照梅军林等[18]的径级划分方法,将不同类型的红背山麻杆划分为:径级Ⅰ(地径≤0.5 cm)、径级Ⅱ(0.5 cm<地径≤1.0 cm)、径级Ⅲ(地径>1.0 cm)。
1.3 研究方法
在2种繁殖类型的红背山麻杆群落中,随机选取35株实生苗,连同其分株一起挖出(包括其根系),观察根系结构,测量红背山麻杆每一基株、根的走向和角度、分株高度和基茎、繁殖根长度和直径、节间长等形态参数。如测定母株与分株、分株与分株之间繁殖根(目的分株与母株相连的根称为前端水平侧根,与另一分株连接的称为后端水平侧根)的长度,用游标卡尺(FH-150,锡丰)测定该部分横走侧根的根径大小;测定每一级侧根与主根的分枝角度,分析每一级横走侧根水平扩展的特点。记录好每一级分株和母株的胸径、冠幅、株高等数据。
1.4 数据统计分析
采用 Microsoft Excel 2010 和IBM SPSS Statistics 22进行统计分析,采用单因素方差分析比较。
2 结果与分析
2.1 红背山麻杆克隆生长构型
分析野外调查结果发现,红背山麻杆属于根源型克隆植物,根系主要分为繁殖根和非繁殖根。繁殖根中细根少,主要是资源的储藏和植株的克隆繁殖;非繁殖根细根较多,主要是吸收土壤水分和营养物质维持植株的生长。桂林喀斯特生境土层浅薄,成土速度极慢,导致部分繁殖侧根裸露在外。研究区红背山麻杆克隆生长的显著特征为:1)分株间的间隔较长,有的可达1 m,分株通常较分散;2)分枝强度较弱,方向性较强,通常一个母株1~2条繁殖水平侧根,少有3条以上;3)分枝夹角一般为60°或50°,为“游击型”克隆生长构型(图1)。

图1 红背山麻杆克隆生长示意Fig. 1 Clonal growth diagram of Alchornea trewioides
2.2 地径与分株数的关系
由图2可知,砍伐干扰克隆繁殖和无干扰克隆繁殖的红背山麻杆分株数均随着地径的增大而增大,复相关系数分别为0.564 8和0.592 5。砍伐干扰克隆繁殖类型斜率为3.583 3,无干扰克隆繁殖类型的斜率为2.549 9,砍伐干扰克隆繁殖类型受地径的影响大于无干扰克隆繁殖类型,说明地径相同时,砍伐干扰克隆繁殖红背山麻杆的克隆繁殖分株数较无干扰克隆繁殖的多。
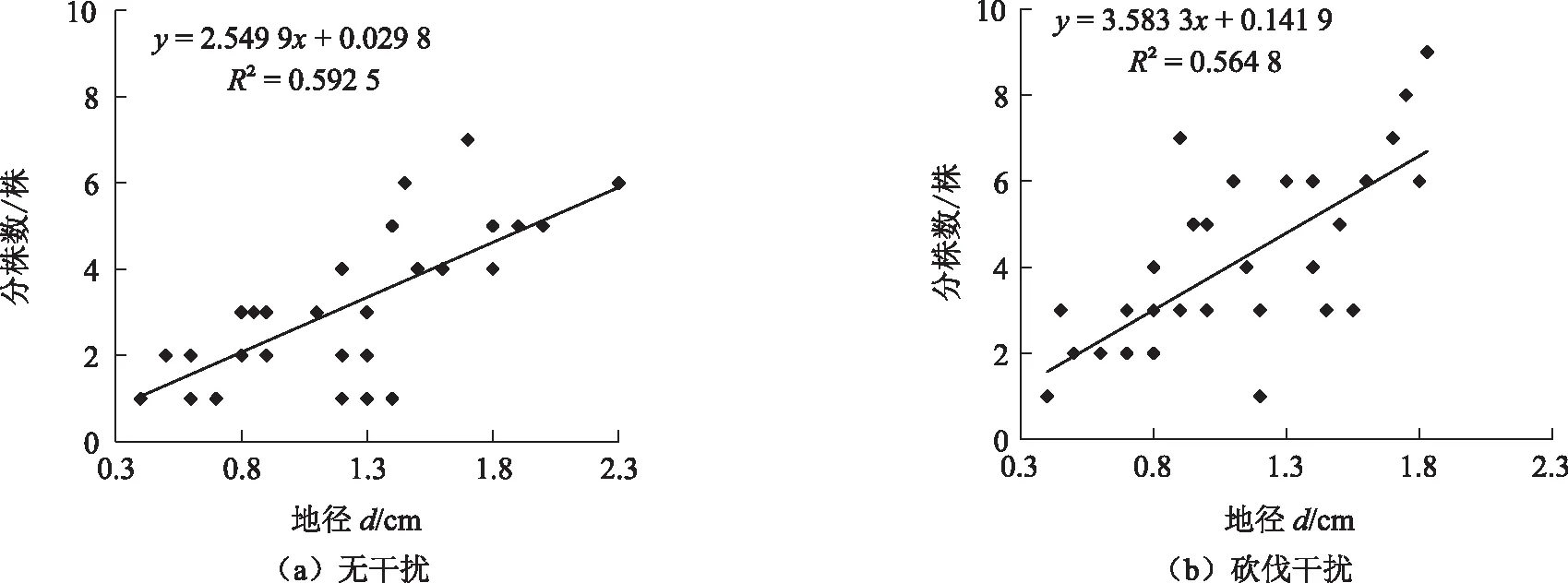
图2 2种类型红背山麻杆地径与分株数的关系Fig. 2 Relationship between basal diameter and number of ramets of two types in Alchornea trewioides
2.3 分株地径与前后端水平侧根直径的关系
由图3和图4可知,无论是砍伐干扰还是无干扰克隆类型,红背山麻杆克隆分株前后端水平侧根直径随着克隆分株地径的增加呈增大趋势。砍伐干扰克隆繁殖类型的地径与前、后端水平侧根直径的复相关系数分别为0.217 1和0.460 8,无干扰克隆繁殖类型的地径与前、后端水平侧根直径的复相关系数分别为0.060 3和0.310 4。砍伐干扰克隆繁殖类型地径与前端水平侧根直径关系的斜率为0.544 5,与后端水平侧根直径的斜率为1.061 1;无干扰克隆繁殖类型地径与前端水平侧根直径的斜率为0.343 5,与后端水平侧根直径的斜率为0.857 7。无论是前端水平侧根直径还是后端水平侧根直径,砍伐干扰克隆繁殖类型受地径的影响均大于无干扰克隆繁殖类型,而后端水平侧根较前端水平侧根更具有线性关系。
2种类型红背山麻杆均随着克隆分株地径的增长,后端水平侧根直径与前端水平侧根直径的差值逐渐增大(图5),说明较小的红背山麻杆分株靠母株供给,随着分株地径的增大,克隆分株能够自行脱离母株供给,独立生存。而分株光合产物用于根茎和分株后端水平侧根的生长多于分株前端水平侧根的生长,所以前端水平侧根直径较小,而后端水平侧根直径较大,并且随着地径的增大,这种前后端水平侧根不对称性逐渐加强。
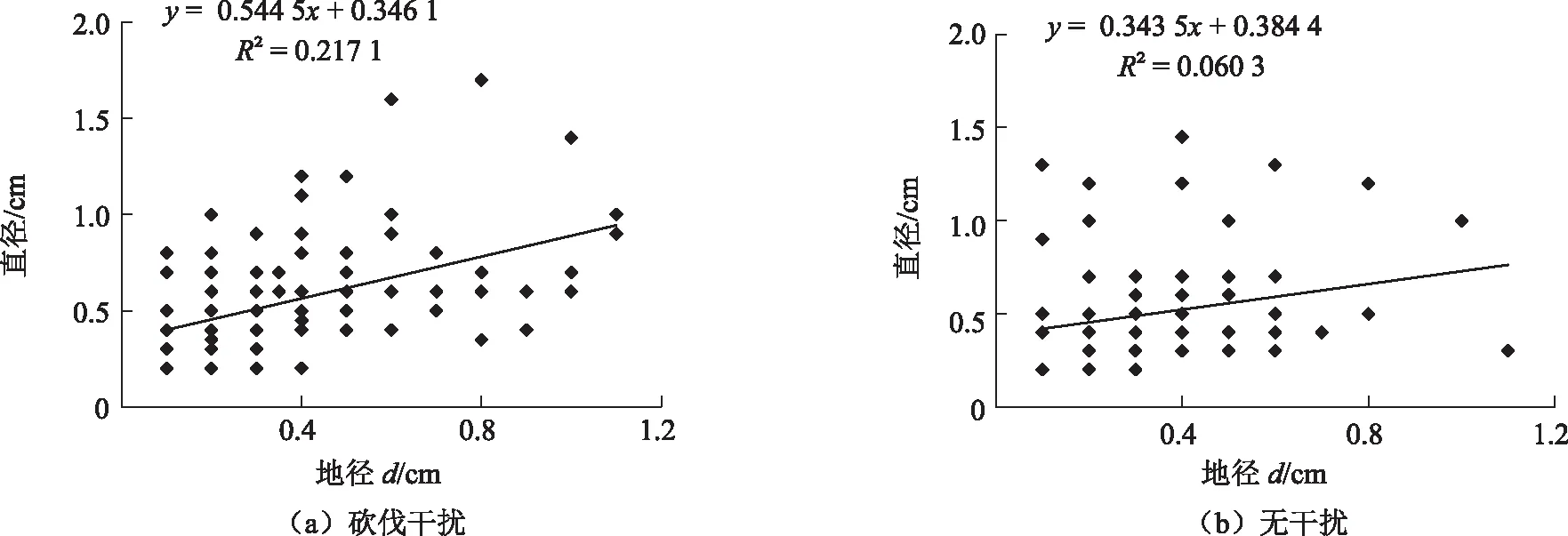
图3 克隆分株地径与前端水平侧根直径的关系Fig. 3 Relationship between base stem diameter and front part of lateral root diameter

图4 克隆分株地径与后端水平侧根直径的关系Fig. 4 Relationship between base stem diameter and back part of lateral root diameter

图5 克隆分株地径与前后端水平侧根直径差的关系Fig. 5 Relationship between base stem diameter and diameter difference of horizontal lateral root at front and rear ends
2.4 不同物候期地径与分株数的关系
由图6可知,无论在哪个物候期,2种类型红背山麻杆径级Ⅲ的分株数均最多。对于径级Ⅰ而言,红背山麻杆克隆繁殖分株数均为果期大于花期和落叶期;对于径级Ⅱ而言,克隆繁殖分株数均为花期大于果期大于落叶期,其中砍伐干扰花期和落叶期达到显著水平;对于径级Ⅲ而言,砍伐干扰红背山麻杆克隆繁殖分株数表现为花期大于果期大于落叶期,其中花期、果期和落叶期达到显著水平,而无干扰红背山麻杆克隆繁殖分株数在3个物候期间保持不变。相同物候期同一径级的红背山麻杆克隆繁殖分株数均为砍伐干扰大于无干扰类型,表明砍伐干扰能够整体促进红背山麻杆的克隆繁殖,增加克隆繁殖强度。
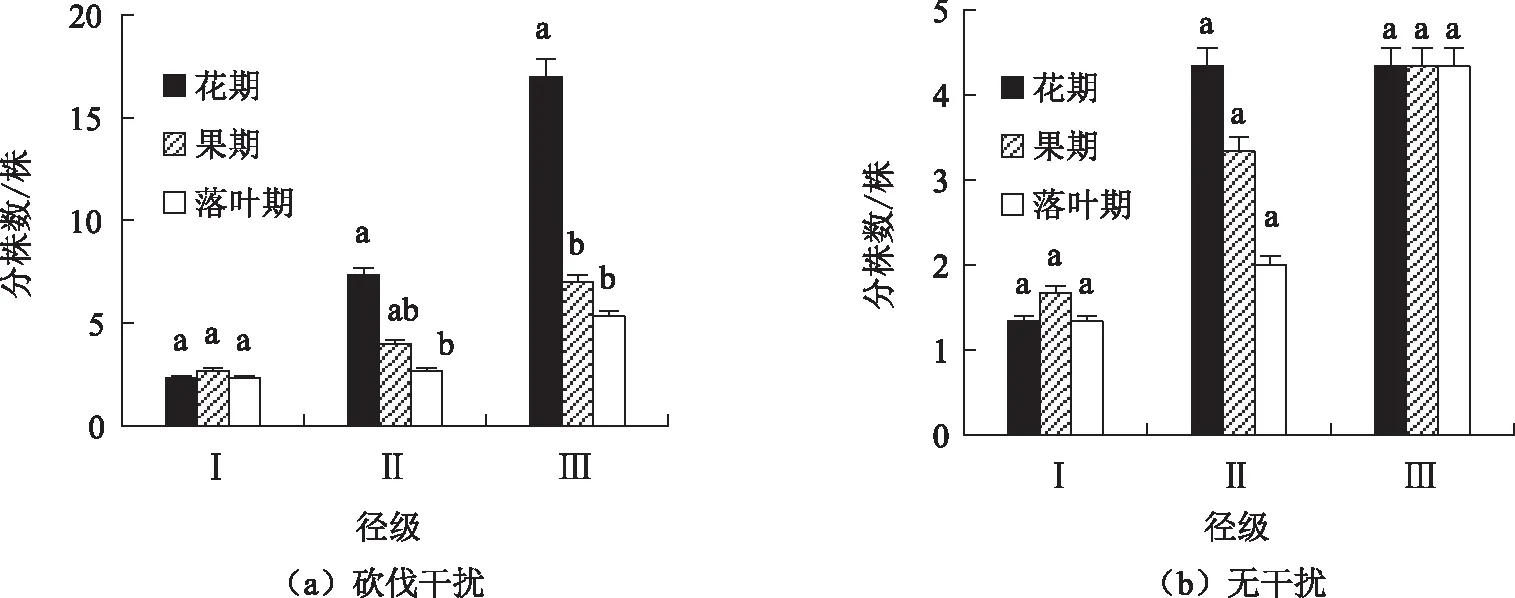
不同小写字母表示不同时期红背山麻杆间差异显著,同一字母表示不存在显著差异图6 3个径级不同物候期的克隆分株数Fig. 6 Number of cloned ramets in different phenological stages of three diameter classes
3 讨论与结论
克隆植物生长型主要是由遗传结构和环境决定的,生长型因具体生境和个体发育阶段不同而变化,不同种群具有不同的生长型[19]。“游击型”构型植物具有很强的克隆扩散能力,易于利用分散分布的资源;而“密集型”构型植物的克隆扩散能力有限,易于利用局部分布的资源[20]。红背山麻杆分枝强度较弱,且方向性较强,分株不密集,表现为“游击型”,这是红背山麻杆对桂林喀斯特生境适应的一种策略,即根据可利用的资源水平差异,通过游击型生长逃避不利生境,向资源较丰富的生境扩展,使其能更好地利用喀斯特生境异质性分布的资源。
相同径级的红背山麻杆,砍伐干扰克隆繁殖类型比无干扰克隆繁殖类型产生的分株数多,说明砍伐干扰能够促进红背山麻杆的克隆繁殖,这与吕美强等[21]研究刈割也能刺激异针茅Stipaaliena、矮嵩草Kobresiahumilis、粗喙薹草Carexscabrirostris和蕨麻Potentillaanserina等克隆植物,增强其克隆繁殖能力的结果类似。砍伐红背山麻杆地上部分,减弱了其有性生殖,但地下部分储存营养,能够快速克隆繁殖出分株。红背山麻杆能够独自利用所有光照、水分等资源,促使分株快速生长,并积累营养于根部继续进行克隆繁殖,能快速形成红背山麻杆优势物种群落。而无干扰的红背山麻杆需要同时进行有性繁殖和克隆繁殖,以保证植株的最优生长和繁殖,加之地上部分没有被砍伐,母株与分株间存在相互竞争关系,才导致克隆繁殖能力比砍伐干扰弱。
2种类型红背山麻杆克隆分株地径与后端水平侧根直径的线性关系均要强于克隆分株地径与前端水平侧根直径的线性关系,这可能是由于前端水平侧根与母株直接相连,受到离母株远近以及母株大小的影响较多,而后端水平侧根则是直接与克隆分株相连,其大小受分株大小影响更多,所以更易表现出与分株地径的线性关系。前后端水平侧根的不对称性与张明如等[22]研究得出火炬树前端水平侧根直径大于后端水平侧根直径的结果相反。本研究表明,随着红背山麻杆克隆分株地径的增大,后端水平侧根直径由一开始比前端水平侧根直径小,越过临界点克隆分株地径0.2 cm后(图5),后端水平侧根直径大于前端水平侧根直径。有研究表明,这种前后端水平侧根直径大小的不对称性应该视为光合有机产物积累不平衡的结果[23-25]。由于喀斯特地区地质条件恶劣、岩石裸露率高、土壤贫瘠,在一般情况下地径小于0.2 cm的克隆分株还没有形成自己的根系,可能还不能独立生存,必须通过母株的营养供给才能生长。前端水平侧根其实为母株的根,根直径随着根系的延伸而变小;随着克隆分株长大,分株具有自己完整的根系,能够独立进行光合作用生存,并且将光合有机产物积累在根部,不同的是红背山麻杆克隆分株具有光合有机产物在水平侧根趋后贮藏的特征,能够将营养物质储藏在根系,提供生长所需的营养物质,以便为其繁殖或者度过恶劣环境做准备,以适应喀斯特环境。水平侧根可作为芽库进行克隆繁殖,也可作为有机营养物质的贮藏库,为红背山麻杆不定芽萌动、分株初期快速生长提供充足的营养物质[24],所以随着其分株的增大,后端水平侧根直径因为光合有机物的积累会比前端水平侧根大,并且这种差距会越来越大。
叶学华[25]研究表明,赖草Leymussecalinus利用克隆繁殖快速产生新的子代分株,分担了基株风蚀死亡风险,使得赖草对风蚀具有较强的抵抗能力。本研究中的砍伐干扰红背山麻杆与此研究结果类似。砍伐干扰红背山麻杆类型的径级Ⅱ、Ⅲ和无干扰红背山麻杆类型的径级Ⅱ克隆繁殖分株数均为花期大于果期大于落叶期,说明此时红背山麻杆克隆繁殖符合风险分摊原理,即,将死亡风险分摊到各分株间,从而保证克隆基株的存活;而无干扰红背山麻杆类型的径级Ⅲ克隆繁殖分株数从果期到落叶期一直保持不变,可能是此时红背山麻杆的繁殖投资达到平衡,在保证有性繁殖的同时也可以维持克隆繁殖分株存活。总体上看,砍伐干扰克隆繁殖的风险分摊现象比无干扰明显,这也间接解释了为什么同径级砍伐干扰克隆分株比无干扰克隆分株多。
