长江口汛期凤鲚捕捞量年际变化研究
2020-06-10倪健夫郭弘艺唐文乔张旭光
倪健夫 ,郭弘艺,唐文乔,,张 亚,张旭光
(1. 上海海洋大学海洋动物系统分类与进化上海高校重点实验室,上海 201306; 2. 上海海洋大学科技部海洋生物学国际联合研究中心,上海 201306; 3. 上海海洋大学水产科学国家级实验教学示范中心,上海 201306)
凤鲚(Coiliamystus)俗称凤尾鱼、烤籽鱼,隶属于鲱形目(Clupeiformes),鳀科(Engraulidae),在我国沿海均有分布,被划分为长江、闽江和珠江3个生态型群体,是一种短距离溯河洄游鱼类[1-2]。长江口是我国凤鲚渔业的主要产区,1960—1998年凤鲚年均产量为1 192 t[3],在20世纪80年代,产量更是高达长江口渔获物总量的48.6%[4]。自20世纪90年代开始,随着长江中下游流域社会经济的快速发展、水域污染的加剧及对资源的长期过度捕捞,凤鲚捕捞量迅速下滑,生物学参数和怀卵量等繁殖特征均发生了显著改变[5-8]。而2003年后三峡工程的蓄水,改变了长江入海径流和泥沙输运原有的季节与年际变化格局[9]。河口环境因子的改变,势必影响到生态系统中生物群落的分布格局和渔业资源的适应性,凤鲚捕捞量呈现近一步大幅下降[10]。近年来,已有学者对凤鲚的资源变动[6-8, 10-11]、摄食习性[12-13]、生活史[14]和种群鉴别[15-19]及脂肪含量等生化成分[20-21]进行了研究报道,但未见关于长江口凤鲚捕捞量格局转变的研究。格局转变贯序t检验法(sequential t-test testing analysis of regime shifts,STARS)可探测稳态改变的发生时间、次数、持续时间及各稳态阶段均值,较传统的Mann-Kendall检验和滑动t检验,对时间序列较短的数据检验更为严谨和准确[22-23],目前已应用于海洋学、生态学、地质与气候科学、环境科学、渔业资源等领域[22-29]。本文利用1982—2014年长江口凤鲚捕捞量数据,结合捕捞船数及环境因子水温、径流和输沙量进行年际格局转变研究,找出格局转变年份。并结合广义可加模型(generalized additive model,GAM)定量分析了凤鲚捕捞量与捕捞船数及环境因子的相关性,旨在弄清长江口凤鲚捕捞量格局转变的影响因子,为凤鲚资源的保护及可持续利用提供基础数据。
1 材料与方法
1.1 数据来源
本研究中,1982—2014年长江口凤鲚汛期(4—6月)的捕捞船数(艘·年-1)、汛期总捕捞量(t·年-1)、单船全汛捕捞量(kg·年-1)数据来源于上海市渔政监督管理处。选取距离长江口最近的长江干流水文监测站大通站的年输沙量(亿t·年-1)和径流量(亿m3·年-1)数据用于分析输沙量和径流量对凤鲚汛期捕捞量的影响,大通站1982—2014年年输沙量和径流量数据来源于水利部长江水利委员会发布的长江泥沙公报[30]。长江口水温(℃)数据来源于东北亚海洋观测系统(NEAR-GOOS)海洋预报系统网[31],网站提供时间跨度为1982年1月—2014年12月长江口(30°40′~31°40′N、 122°00′~122°20′E)海表水温,分辨率为月。本研究选取凤鲚捕捞期4—6月水温数据。
1.2 数据处理和模型建立
采用Origin 8.6软件对数据进行处理和绘图,并用互相关函数(cross-correlation function)对凤鲚捕捞量和环境因子的水温、径流和输沙量进行互相关分析(cross-correlation analysis)。
采用格局转变贯序t检验(STARS)的方法计算凤鲚捕捞量、捕捞船数和环境因子水温、径流量、输沙量的年际格局转变指数(regime shift index,RSI)。t0时刻的RSI值公式为[32]:
RSI=ttest(t0)=
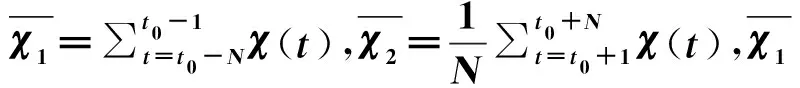
凤鲚捕捞量与捕捞船数及环境因子间的关系进一步采用广义可加模型(GAM)拟合,将凤鲚总捕捞量和单船全汛捕捞量分别作为模型响应变量,以捕捞船数、水温、径流和输沙量作为解释变量建模。由于4—6月水温之间互相显著相关,会造成模型多重共线性,同时考虑到5月为主汛期,月捕捞量比例高达61.2%[10],因此,选用5月水温数据作为解释因子用于建模。本研究中,采用逐步回归的方法筛选模型变量,即将解释变量逐一输入到模型中,根据F检验评估变量的显著性,若P<0.05时,保留此变量。依次增加变量,最后根据模型赤池信息量准则(Akaike information criterion,AIC)越小即表明模型拟合效果越佳的原则[33-34],确定最终模型。GAM模型拟合和绘图均采用R 3.1.1软件中的mgcv包实现[33]。
2 结果与分析
2.1 捕捞船数和捕捞量的年际格局转变
研究结果显示,1982—2014年长江口凤鲚汛期捕捞船数变幅为59~330艘·年-1,表现为先增后减,总体呈下降趋势。而总捕捞量和单船全汛捕捞量变幅分别为7.9~3 252.0 t·年-1和130.0~12 945.5 kg·年-1,均呈下降趋势。利用STARS研究1982—2014年中长江口凤鲚汛期捕捞船数、总捕捞量和单船全汛捕捞量格局转变的年份,分析转变前后特征变化(图1~图3)。
结果显示,捕捞船数分别在1990年、2000年和2006年发生3次格局转变,对应RSI值分别为0.992、-0.412和-0.763。在1990年格局转变前,捕捞船数保持在平均191艘·年-1,1990年后捕捞船数大幅增加,上升至平均298艘·年-1,增幅达55.5%;2000年之后捕捞船数开始下降,平均为207艘·年-1;至2006年捕捞船数进一步大幅减少,平均为86艘·年-1。
汛期总捕捞量分别在1997年和2007年发生2次格局转变,对应RSI值分别为-0.580和-0.405,即2次转变均为下降态势。1997年格局转变之前总捕捞量较高,平均高达1 844.15t·年-1,1997年之后总捕捞量降低,平均为863.88 t·年-1,较格局转变前下降了53.2%;至2007年第二次格局转变后,总捕捞量锐减至75.6 t·年-1,几不成汛。
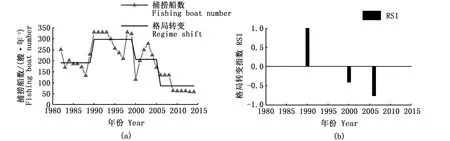
图1 长江口1982—2014年凤鲚汛期捕捞船数的年际格局转变(a)和格局转变指数(b)Fig.1 Annual variations, regime shift (a) and regime shift index (b)of Coilia mystus fishing boat numbers in the Yangtze River Estuary during 1982—2014

图2 长江口1982—2014年凤鲚汛期总捕捞量的年际格局转变(a)和格局转变指数(b)Fig.2 Annual variations, regime shift (a) and regime shift index (b)of Coilia mystus total catch in the Yangtze River Estuary in fishing season during 1982—2014

图3 长江口1982—2014年凤鲚汛期单船全汛捕捞量的年际格局转变(a)和格局转变指数(b)Fig.3 Annual variations, regime shift (a) and regime shift index (b)of Coilia mystus CPUE in the Yangtze River Estuary in fishing season during 1982—2014
1982—2014年中,凤鲚单船全汛捕捞量亦经历2次下降的年际格局转变,转变年份为1990年和2007年,对应RSI值分别为-0.806和-0.371。1990年之前,单船全汛捕捞量平均为10 460.1 kg·年-1;格局转变后减少至4 711.6 kg·年-1,下降了55.0%;到2007年后进一步减少为720.8 kg·年-1,降幅达84.7%(图3)。
综上可见,在1990年年际格局转变后,单船全汛捕捞量下降了55.0%,相应捕捞船数提高了55.5%,延迟了总捕捞量的下降,甚至在1995年出现了昙花一现的总捕捞量峰值,但至1997年后总捕捞量大幅下滑。捕捞强度的提高,仅短期提升总捕捞量。至2006—2007年格局转变后,汛期捕捞船数、总捕捞量和单船全汛捕捞量均大幅减少。
2.2 环境因子影响
2.2.1 长江口水温
由图4可知,长江口4月、5月和6月水温在1982—2014年33年内总体均呈增高趋势。水温在1997格局转变,RSI系数分别为0.465、0.498和0.404。4月、5月和6月平均水温分别为11.3℃、13.7℃和17.5℃;1997年后,4月、5月和6月平均水温分别增至13.3℃、16.0℃和19.7℃。互相关分析结果显示,4月、5月和6月长江口水温均与凤鲚总捕捞量及单船全汛捕捞量呈显著负相关关系(P<0.05)(图5,图6)。

图4 长江口1982—2014年4—6月水温的年际格局转变(a)和格局转变指数(b)Fig.4 Annual variations, regime shift (a) and regime shift index (b)of the surface water temperature in the Yangtze River Estuary from April to June during 1982—2014
2.2.2 径流量和输沙量
1982—2014年大通站径流量和输沙量变化范围分别为6 671~12 490亿m3·年-1和0.72~5.05亿t·年-1,总体均呈下降趋势。由图7、图8可见,径流量和输沙量均在2006年发生下降的年际格局转变,对应RSI分别为-0.110和-0.862。格局转变前,平均径流量和输沙量分别为9 217亿m3·年-1和3.44亿t·年-1,2006年后,降低为8 268亿m3·年-1和1.24亿t·年-1,较之前分别下降10.4%和63.9%。
互相关分析结果显示,径流量与凤鲚总捕捞量、单船全汛捕捞量均无显著相关关系(P>0.05)(图9)。但输沙量与凤鲚总捕捞量、单船全汛捕捞量均呈显著正相关关系(P<0.05),且时滞年份为0时,正相关系数最大(图10)。

图5 长江口1982—2014年凤鲚汛期总捕捞量与长江口4—6月水温的互相关分析Fig.5 Cross-correlation analysis between Coilia mystus total catch in fishing season and surface water temperature in the Yangtze River Estuary from April to June during 1982—2014注:虚线代表P=0.05显著水平Note: The dashed line indicates the significance level (P=0.05)

图6 长江口1982—2014年凤鲚单船全汛捕捞量与长江口4—6月水温的互相关分析Fig.6 Cross-correlation analysis between Coilia mystus CPUE and surface water temperature in the Yangtze River Estuary from April to June during 1982—2014注:虚线代表P=0.05显著水平Note: The dashed line indicates the significance level (P=0.05)
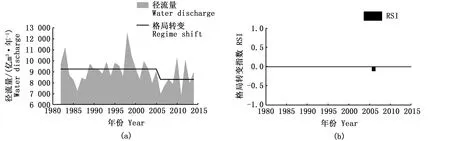
图7 长江大通水文站1982—2014年径流量年际格局转变(a)和格局转变指数(b)Fig.7 Annual variations, regime shift(a) and regime shift index (b)of water discharge at Datong Hydrological Station during 1982—2014

图8 长江大通水文站1982—2014年输沙量年际格局转变(a)和格局转变指数(b)Fig.8 Annual variations, regime shift (a) and regime shift index (b)of sediment discharge at Datong Hydrological Station during 1982—2014

图9 长江口1982—2014年凤鲚汛期总捕捞量(a)和单船全汛捕捞量(b)与大通站径流量的互相关分析Fig.9 Cross-correlation analysis between Coilia mystus total catch in fishing season and water discharge (a), and between Coilia mystus CPUE and water discharge (b) at Datong Hydrological Station during 1982—2014注:虚线代表P=0.05显著水平Note: The dashed line indicates the significance level (P=0.05)
2.3 GAM模型结果
GAM模型在构建过程中需要逐步加入各个解释变量及其交互项,根据F检验和AIC值来选择最优模型,GAM模型的最终选择结果见表1和表2。模型对长江口凤鲚汛期总捕捞量和单船全汛捕捞量的总偏差解释率分别为97.4%和90.2%,表明2个模型均较好拟合各解释变量与捕捞量之间的关系。
F检验表明,径流量对凤鲚总捕捞量影响不显著(P>0.05),而捕捞船数、水温和输沙量对凤鲚总捕捞量具有显著影响(P<0.05)(表1)。其中,捕捞船数对凤鲚总捕捞量影响最大,呈非线性正相关关系(图11),偏差解释率高达89.2%,而环境因子中的水温和输沙量对凤鲚总捕捞量亦存在影响,偏差解释率分别为4.9%和3.3%(表1)。
GAM模型还显示,对凤鲚单船全汛捕捞量影响最大的因子为输沙量,偏差解释率高达55.2%,随着输沙量的增加,单船全汛捕捞量随之增大(图12-a)。其次为捕捞船数,偏差解释率为33.5%,二者亦呈非线性的正相关关系。捕捞船数在50~150艘·年-1区间内,单船捕捞量随着捕捞船数的增加而增大;而在捕捞船数超过150艘·年-1后,单船捕捞量随之增加不显著(图12-b)。水温的影响最弱,偏差解释率为1.5%(表2)。水温与凤鲚单船全汛捕捞量呈线性负相关,随着水温升高,单船全汛捕捞量随之降低(图12-c)。
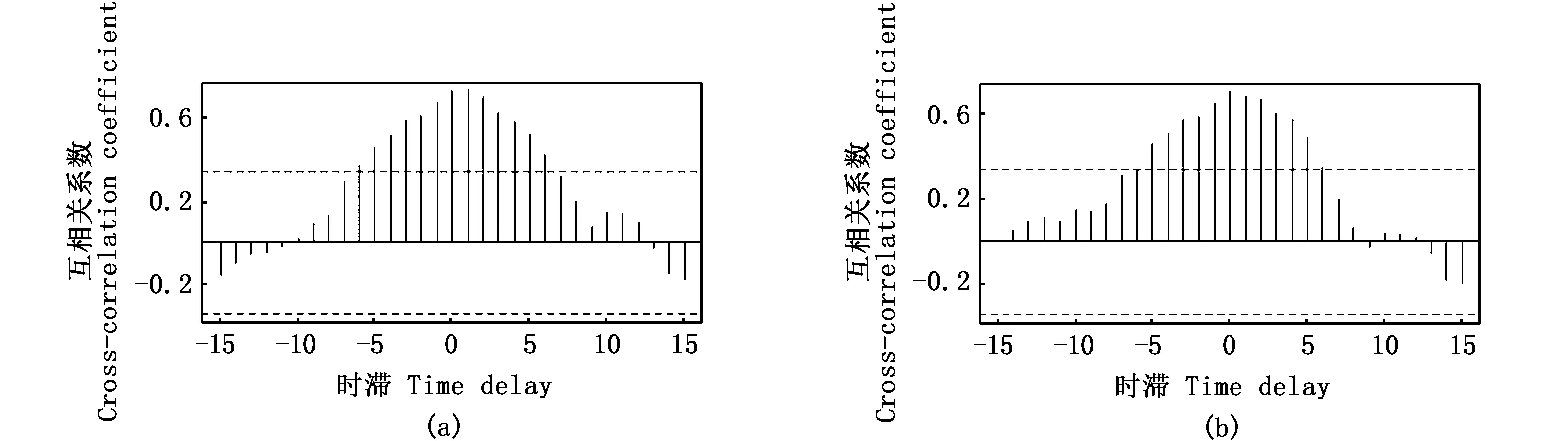
图10 长江口1982—2014年凤鲚汛期总捕捞量(a)和单船全汛捕捞量(b)与大通站输沙量的互相关分析Fig.10 Cross-correlation analysis between Coilia mystus total catch in fishing season and sediment discharge (a),and between Coilia mystus CPUE and sediment discharge (b) at Datong Hydrological Station during 1982—2014注:虚线代表P=0.05显著水平Note: The dashed line indicates the significance level (P=0.05)
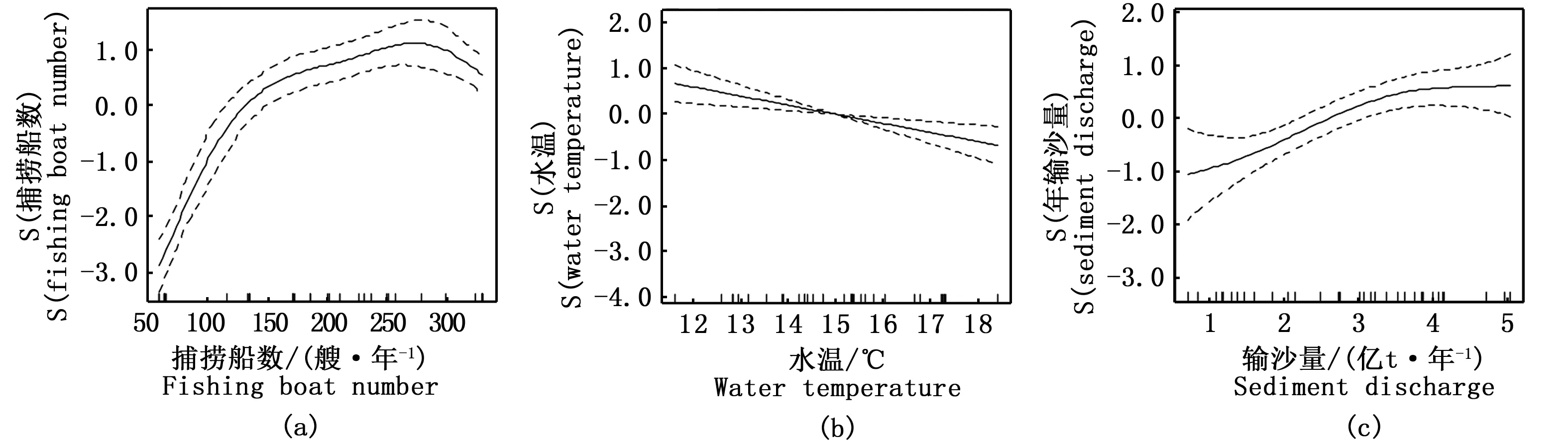
图11 广义可加模型(GAM)所揭示的显著影响长江口凤鲚汛期总捕捞量变化的因子:(a)捕捞船数, (b)水温, (c)输沙量 Fig.11 Generalized additive model (GAM) derived effects of fishing boat number(a), water temperature(b), sediment discharge(c), on total catch of Coilia mystus in the Yangtze River Estuary during 1982—2014注:S (因子)为光滑样条函数的拟合值,表示其对凤鲚总捕捞量的影响。实线表示凤鲚总捕捞量的期望值,上下两侧虚线表示方程的95%置信区间Note: S(factor) is the fitted value of smoothing spline functions, representing their impacts on total catch.The solid lines represent the expected values of total catch; the dotted lines represent the 95% confidence intervals of equations

模型因子Model factors残差自由度df残差偏差Residual deviance偏差变化量Deviance variation累计解释偏差/%Accumulation of deviance explainedPAICNull32.0097.22+S(捕捞船数)28.1810.5086.7289.2<0.00166.29+S(水温)22.795.754.7594.10.002 956.47+S(年输沙量)14.812.453.3097.40.012 642.23

表2 长江口凤鲚单船全汛捕捞量与相关因子GAM检验Tab.2 Significance of explanatory variables effects on Coilia mystus CPUE in the Yangtze River Estuary during 1982—2014 as determined by GAM
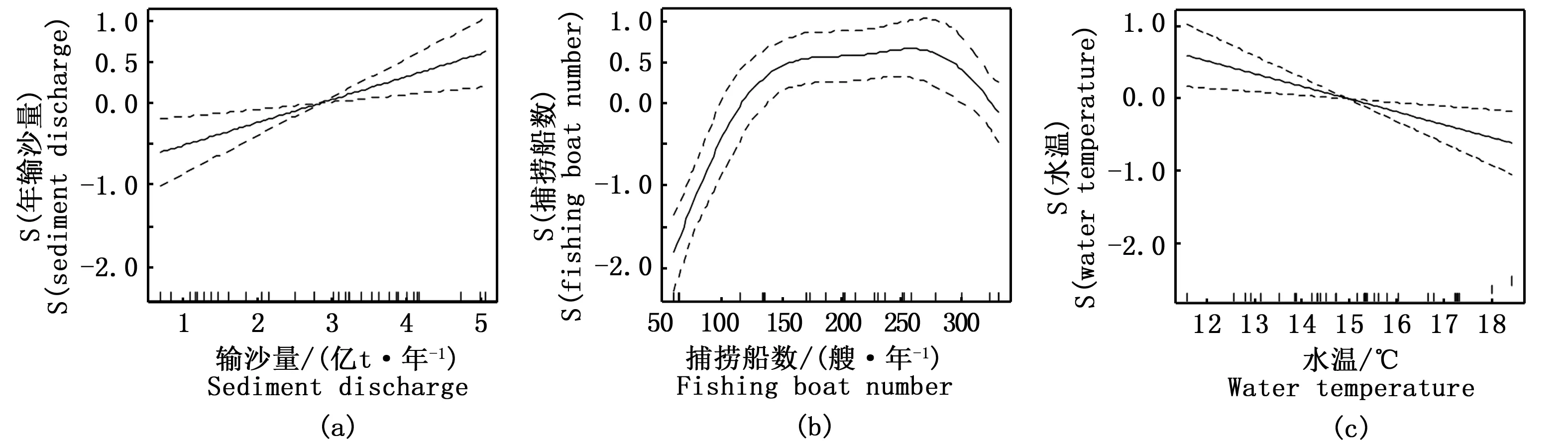
图12 广义可加模型(GAM)所揭示的显著影响长江口凤鲚单船全汛捕捞量变化的因子:(a)年输沙量, (b)捕捞船数, (c)水温Fig.12 Generalized additive model (GAM) derived effects of sediment discharge(a), fishing boat number(b),water temperature(c) on Coilia mystus CPUE in the Yangtze River Estuary during 1982—2014注:S (因子)为光滑样条函数的拟合值,表示其对单船捕捞量的影响。实线表示单船全汛捕捞量的期望值,上下两侧虚线表示方程的95%置信区间Note: S(factor) is the fitted value of smoothing spline functions, representing their impacts on CPUE. The solid lines represent the expected values of CPUE; the dotted lines represent the 95% confidence intervals of equations
3 讨论
1984—2014年长江口凤鲚汛期总捕捞量和单船全汛捕捞量变幅分别为7.9~3 252.0 t·年-1和130.0~12 945.5 kg·年-1,均呈下降趋势。张国祥和华家栋[4]研究指出,自20世纪70年代起长江口凤鲚捕捞强度持续多年强化,使得凤鲚的补充群体逐年减少,至80年代后凤鲚资源量已明显下降。在本研究中,1990年之前,捕捞船数保持在平均191艘·年-1,单船全汛捕捞量平均高达10 460.1 kg·年-1,捕捞量随捕捞船数波动响应明显;1990年后,捕捞船数增至207艘·年-1,单船全汛捕捞量反而减少至4 711.6 kg·年-1,期间仅在1995年出现“昙花一现”的高产,捕捞量对捕捞船数的响应大幅减弱。STARS分析结果显示,作为凤鲚资源量波动指示因子之一的单船全汛捕捞量在1990年和2007年发生3次下降的年际格局转变,其下降即预示凤鲚资源的衰退。究其原因,1990年第一次格局转变可能是由于过度捕捞最终导致凤鲚种群调节机制被破坏,凤鲚种群恢复能力已逐渐无法适应过高的捕捞强度,故而造成资源的衰退。
STARS分析还显示,捕捞船数、大通水文站径流量和输沙量都在2006年发生格局转变,这与凤鲚单船汛期捕捞量的第二次格局转变时间节点基本吻合。长江口素有来水来沙量大的特点,1986年前通过大通水文站输入长江口的平均年输沙总量高达4.64亿t·年-1,受温度、盐度、潮流、径流等因素的影响,长江口门附近形成最大浑浊带,长江口的高悬沙含量及复杂的行为动态在世界各河口也是非常少见的[35]。研究显示,近三十年来,通过大通水文站的输沙量呈波动下降态势,三峡建库后输沙量降低更为显著,2006—2014年长江大通站输沙量年均仅为1.24亿t,较之前下降了63.9%。其中,在不考虑沿程冲刷恢复的条件下,中等粒径颗粒的泥沙减少更为明显[9]。输沙量的减少使得河口及其邻近海岸动力作用和泥沙沉积过程发生变化,不可避免地对长江下游及河口的生态环境产生显著影响,从而影响河口鱼类及浮游生物的空间格局[36-38]。同时,由于入海泥沙量的减少所引起的河口水域海水透明度的增加,可能使得浮游植物高生产力区向海岸扩展,许多鱼类的产卵索饵场位置亦会有相应的变化[39]。GAM模型亦显示,输沙量与凤鲚单船全汛捕捞量存在显著正相关关系,综上推测,长江口输沙量减少所造成的水文条件改变,可能对凤鲚资源产生了负面影响。
研究期间,通过大通站的径流量在2006年后较之前下降10.4%。径流量的降低可能主要受三峡工程蓄水的影响。DRINKWATER 和FRANK[38]曾提出河口径流的变动可引起温度、盐度、浊度和溶解氧的变化,这些效应最终都影响到河口生物的丰度和分布类型。董方勇[40]认为水利工程造成河口区4—7月的径流量减少,使得河口海水上溯的盐度随之发生变化,从而可能造成凤鲚产卵场西移以及生殖洄游时间推迟。管卫兵等[8]研究亦显示,径流量的减少对凤鲚的生长和生殖具有较强的负面影响。刘凯等[10]报道,三峡蓄水后长江口凤鲚生物学参数和汛期繁殖力均呈下降趋势,绝对怀卵量和相对怀卵量较蓄水前分别下降17.38%和16.97%、全长和体质量较蓄水前分别下降4.58%和3.55%。本研究中,GAM模型和互相关分析均显示,径流量对长江口凤鲚捕捞量的影响未达显著水平,但由于径流时间格局的转变至今尚不足10年,因此,仍然需要长期监测和谨慎评估径流量减少对凤鲚乃至其他鱼类的影响。
此外,GAM模型还显示长江口汛期水温亦是影响长江口凤鲚总捕捞量的重要环境因子之一。由于江海洄游型鱼类在生殖洄游过程中一般并不摄食,依靠体内积累的营养物质完成溯河洄游的运动、代谢和生殖等过程,因此在生殖洄游开始前需要积累大量的脂肪[41]。凤鲚这类短途溯河繁殖的洄游鱼类,其卵巢粗脂肪含量达60%以上、肌肉粗脂肪含量达1.92%以上(鲜重),才开始生殖洄游[21]。管卫兵等[8]的研究表明,水温升高年份的凤鲚生殖群体的丰满度和性腺重量反而下降。可能的解释是,水温的升高使得鱼类代谢增强、体内营养物质(粗脂肪等)的积累减缓[42-43],从而影响凤鲚群体的成熟、集群和洄游过程,进而可能影响长江口凤鲚资源量。
除环境因子以外,长江口鳗苗网的兼捕对凤鲚补充群体损害也极为严重[44],自2007年起,长江口凤鲚的鱼卵和仔稚鱼丰度呈现大幅降低趋势[45]。环境因子和捕捞因素的综合影响可能是造成凤鲚单船汛期捕捞量的第二次格局转变的原因。但凤鲚的捕捞量变化不能简单地归因于一种或几种原因,在这一长期变化过程中更多的是生态因素与人为扰动相互作用的结果,关于更多影响因素和影响机制的研究也亟待展开。
