喀斯特不同恢复阶段植物根际土养分和酶活性的季节性变化和根际效应
2020-06-02潘复静王克林梁月明靳振江
潘复静,王克林,张 伟,梁月明,靳振江,李 臻
(1.桂林理工大学 a.环境科学与工程学院;b.广西环境污染控制理论与技术重点实验室,广西 桂林 541006;2.中国科学院亚热带农业生态研究所 亚热带农业生态过程重点实验室,长沙 410125;3.中国科学院环江喀斯特生态系统观测研究站,广西 环江 547100;4.中国地质科学院岩溶地质研究所/自然资源部广西壮族自治区岩溶动力学重点实验室,广西 桂林 541004)
0 引 言
土壤氮磷是生态系统中植物生长不可或缺的元素,而碳也是物质循环和能量流动的重要组成部分。土壤碳氮磷元素的主要来源是土壤有机质和母岩[1]。土壤有机质主要由植物根、茎和叶的枯落物分解产生,经过淋洗、机械破碎、土壤腐食动物啃食和酶解等过程,最终释放碳氮磷元素到土壤中[2];另一方面,植物根系和微生物共同分泌有机酸腐蚀岩石,并通过土壤酶转化元素的形态,从而达到释放元素的目的[3-4]。土壤酶主要由土壤微生物和植物根系分泌产生,是参与土壤碳氮磷养分循环过程的重要物质,能提高矿质元素的有效性[5]。在植物和土壤物质交换的重要场所根际区,矿质元素有效性的提高能保证植物的养分吸收。
植物根际土壤酶活性的提高对植物适应土壤养分缺乏或提高土壤养分有效性非常重要。一般情况下,根际土壤酶的活性与植物物种具有密切关系[6]。 不同植物的根系分泌物、脱落物与植物残体的质量和数量具有很大差异,影响土壤有机物的分解,同时养分吸收效率也不同,最终导致不同植物根际区土壤酶活性的差异。 随着植被演替的正向发展,植物多样性逐渐提高,物种结构组成也呈现复杂化。 在这种情况下,植物对土壤养分的需求量也在增加,而土壤养分的缺乏会阻碍植物对养分的吸收。 在土壤养分缺乏的情况下,为了能从土壤有机质中获取更多的养分元素,植物可以刺激土壤微生物的活力,诱使根际区微生物分泌更多的胞外酶[3]。 土壤酶活性是提高土壤养分有效性的重要影响因素之一。β-葡聚糖酶(βG)和N-乙酰-β-D-葡萄糖苷酶(NAG)是土壤中几种关键的胞外酶,它们的分泌有利于加快碳氮磷等矿质元素从土壤有机质和母质中释放,提高土壤中有机物的降解速率和氮磷等养分的有效性[5]。同时,土壤有机碳含量的提高为微生物生长提供充足的能量,从而提高微生物的数量和活性,进而促进胞外酶的分泌,保证微生物生长和酶分泌两者关系的良好发展。特别是在土壤钙含量较高的地区,植物根系分泌有机酸等有机物质降低土壤有机质的稳定性,使酶和有机物质的可接触面积增多。
中国西南喀斯特地区是世界上最大的连片喀斯特脆弱区,面积为54万km2,土层浅薄且分布不连续、普遍缺乏C层、成土速率慢[7]。20世纪中后期,强烈的人类活动导致喀斯特地区森林植被破坏严重,造成水土流失、土地退化、基岩裸露、石漠化加剧以及生态系统服务结构和功能退化,从而制约了该地区社会、经济、环境的可持续发展[8-10]。近年来,随着大规模石漠化治理和生态恢复工程的实施,喀斯特部分重度、中度石漠化区逐渐演变为中度、轻度石漠化区,石漠化趋势得到了一定程度的遏制[11],但是形势仍然很严峻。植被恢复是喀斯特地区石漠化治理的最有效措施之一。在喀斯特地区,植被恢复后期的年第一生产力和生物量均高于恢复前期[12-13],但是植被恢复后期的土壤肥力上升速率比植被年第一生产力的低[14-15],最终造成植物生长受到了土壤养分缺乏的限制。不同植被恢复阶段植物生长受土壤养分缺乏限制不同:植被恢复前期植物生长受限于土壤氮素缺乏,而恢复后期则受限于土壤磷素缺乏[16]。
在喀斯特地区,关于土壤酶活性的研究多集中在植被恢复阶段、地形、树种和土壤理化性质对非根际土壤酶活性的影响[17-20],少数涉及这些因素对根际土壤养分和酶活性的影响[21-22],但是较少关注不同季节、不同植物根际土壤酶活性的变化与土壤养分有效性的关系。因此,本文将研究不同季节、不同植被恢复阶段的6种不同植物的土壤养分和胞外酶活性的变化,通过比较根际土和非根际土的土壤养分和胞外酶活性的差异研究不同季节、不同植物的根际效应特征,揭示土壤养分限制差异情况下植物根际环境的变化和机理,为本地区生态恢复和石漠化治理方式提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区包括广西河池市环江毛南族自治县的中国科学院环江喀斯特生态系统观测研究站(24°43′58.9″—24°44′48.8″ N,108°18′56.9″—108°19′58.4″ E)和木论国家级自然保护区(25°06′09″—25°12′25″ N,107°53′29″—108°05′42″ E)。该地区为典型亚热带季风气候,年均温度16.5 ~20.5 ℃,年均降水量1 400 ~ 1 500 mm,雨季平均持续 130 ~ 140 d,主要集中在4—9月,但降雨时间分布不均,以6月中旬至7月中旬最多,洼地常涝,10月至次年3月为旱季[23]。日照时数以2月份最低,7月份最高;日照率以3月份最低,9月份最高[24]。两个研究点均属于典型喀斯特峰丛洼地景观,土壤pH值为6.3~7.9,洼地和坡面土壤深度分别为50~ 80 cm 和10 ~ 30 cm。
中国科学院环江喀斯特生态系统观测研究站所在地的植被在1958年至20世纪80年代严重退化,从1985年左右开始退耕,至今植被自然恢复已达30年。该研究点70 % 左右的面积被灌丛覆盖,灌丛植物优势物种主要有红背山麻杆(Alchorneatrewioides)、灰毛浆果楝(Cipadessacinerascens)、深紫木蓝(Indigoferaatropurpurea)、小蜡(Ligustrumsinense)、牡荆(Vitexnegundo)、盐肤木(Rhuschinensis)等[25]。
环江木论国家级自然保护区建于1991年,距离环江喀斯特生态系统观测研究站约35 km,总面积为108.6 km2,森林覆盖率93.4 %,生长着中亚热带隐域性的石灰岩森林原生植被,以喀斯特森林生态系统为主要保护对象[26]。保护区的原生林已有200年未受到人类干扰,其优势植物物种主要有紫弹树(Celtisbiondii)、灰岩棒柄花(Cleidionbracteosum)、厚壳桂(Cryptocaryachinensis)、青冈栎(Cyclobalanopsisglauca)、檵木(Loropetalumchinens)、野独活(Miliusachunii)、青檀(Pteroceltistatarinowii)等[27-28]。
1.2 土壤样品采集
在灌丛阶段选取红背山麻杆(A.trewioides)、小蜡(L.sinense)和深紫木蓝(I.atropurpurea)3种优势灌木,在原生林阶段选取檵木(L.chinense)、紫弹树(C.biondii)、青檀(P.tatarinowii)3种优势乔木,分别在2014年1月(旱季中期)和7月(雨季中期)采集根际土和非根际土样品。 为了消除坡位对各指标测试结果可能存在的影响,两种不同植被类型样地中均设置了3个坡位(下坡、中坡和上坡)的样方[29]。为了减少植物物种和个体之间的相互效应,每个坡位选取的灌木物种个体之间的距离不小于50 cm,乔木物种个体之间的距离不小于10 m。
根际土是指抖落松散土之后仍然粘附在根系表面的土,其余的则为非根际土[30]。 根际土采集步骤如下: 把土壤0 ~15 cm深度的细根和土壤挖掘出来,轻轻抖动细根和土壤使松散的土掉落,轻取细根表面的土壤[31], 同时采集非根际土。 本研究分别采集到根际土和非根际土样品各132个(ie: 6种植物×3个坡位×2个季节×4(如不够4个,则取3个)重复),共264个土壤样品。
土壤样品装入塑料袋,放入冰盒中保存,迅速带回实验室进行处理。土壤样品中的细根碎屑在样品采集4 h内全部挑出,然后将土样进行多次混匀,过10目(2 mm)筛,并分成等量的两份:第一份立即放入-20 ℃冰箱保存,用于土壤胞外酶活性的测定;第二份风干,研磨,过20目(0.85 mm)和100目(0.15 mm)筛,用于土壤养分含量的测定。
1.3 样品分析
1.3.1βG和NAG酶活性测定 土壤βG酶活性底物为4-甲基伞形酮酰-β-D-吡喃葡糖酸苷(4-MUB-β-D-glucoside),NAG则为4-甲基伞形酮酰-β-D-吡喃葡糖酸苷(4-MUB-N-acetyl-b-D-glucosamin-ide)。称取1 g土样并置于125 mL 50 mmol/L,pH 5.0 醋酸缓冲液中,用高速搅拌器匀浆1 min制成悬浊液。悬浊液用磁力搅拌器保持匀浆状态,等分成8份,每份以200 μL转移到96格微孔板的小格中。50 μL 200 μmol/L的底物加入到每个小格中。每一块微孔板都有8个空白、阴性对照和标准样。所有样品和溶液全部准备好之后,微孔板则放入20 ℃温室中黑暗状态下培养2 h。培养结束之后,每个小格中滴入10 μL 1.0 mol/L NaOH溶液终止反应。βG和NAG酶活性则用酶标仪(Infinite M200 PRO; TECAN,Switzerland)在365 nm光谱下激发和450 nm光谱下检测测定4-甲基伞形酮(4-MUB,4-methylumbelliferyl)的含量(μmol·g-1soil·h-1)[32]。
1.3.2 土壤化学特征测定 土壤有机碳含量(SOC)采用KCr2O7+H2SO4氧化法测定; 全氮(TN)采用凯氏定氮法并用流动注射仪(FIAstar 5000,FOSS,Hillerød,Denmark)测定; 全磷(TP)采用加NaOH后放入马弗炉高温消煮,H2SO4+HCl清洗后以钼蓝显色液进行显色,用分光光度计进行测定;土壤有效氮(AN)采用碱解扩散法测定;土壤有效磷(AP)用NaHCO3溶液浸提后以钼蓝显色液显色,用分光光度计进行测定[28]。
1.4 统计与分析
用Excel进行数据整理,SPSS 11.5对土壤SOC、N和P 全量和有效含量及β-葡聚糖酶(βG)和N-乙酰-β-D-葡萄糖苷酶(NAG)活性进行正态性检验,均服从正态分布(p>0.05)。 用多重比较分析方法(LSD)对不同植物物种的根际土和非根际土土壤养分含量及土壤酶活性的差异进行了分析。 根际效应(R/B值:R表示根际土的各项指标的值,B表示非根际土的各项指标的值)采用根际土各指标除以非根际土对应的指标,比值越大根际效应越明显[33]。用Pearson方法分析了土壤养分含量与酶活性的相关性。
2 结果与分析
2.1 土壤养分的季节变化及其根际效应
表1为喀斯特植物根际/非根际土样各类形态碳氮磷养分含量及酶活性平均值统计分析。旱季,除了檵木的根际土有机碳(SOC)显著高于非根际土之外,其余树种无显著差异。 6种植物的根际土全氮含量(TN)显著高于非根际土,但是根际土和非根际土的全磷(TP)含量无显著差异。 除了小蜡和檵木的根际土有效氮(AN)含量显著高于非根际土之外,其余无显著差异。 除了红背山麻杆、小蜡和青檀的根际土有效磷(AP)含量显著高于非根际土之外,其余树种无显著差异。雨季,6种植物的根际土和非根际土的SOC和TP含量无显著差异;红背山麻杆、小蜡、深紫木蓝和紫弹树的根际土TN含量显著高于非根际土; 红背山麻杆、小蜡、深紫木蓝和檵木的根际土AN含量显著高于非根际土; 红背山麻杆、小蜡和深紫木蓝的根际土AP显著高于非根际土。
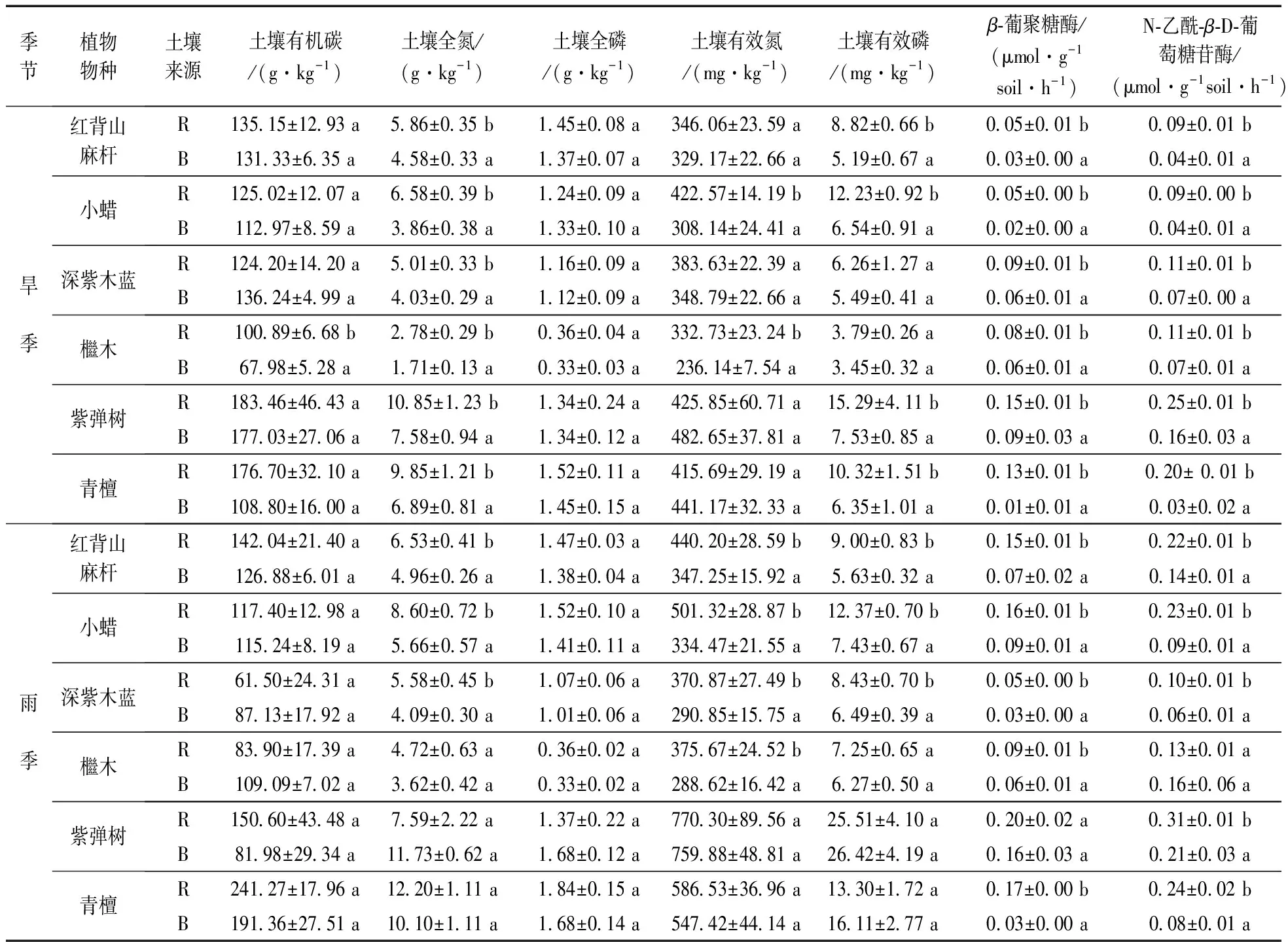
表1 喀斯特植物根际土(R)和非根际土(B)碳氮磷含量和有效性及酶活性Table 1 Nutrients concentrations and availabilities in rhizosphere(R) and bulk(B) soils of six plants in karst ecosystem
注: a和b代表同一植物的非根际土和根际土具有显著差异(p< 0.05)。
灌木阶段和原生林阶段的根际土和非根际土AN和AP含量在雨季大于旱季,但是SOC、TN和TP的季节性变化不明显。
无论是旱季还是雨季,灌木阶段和原生林阶段植物物种的AN和AP均出现根际效应,但是TP未出现根际效应,SOC的根际效应不够明显;红背山麻杆、小蜡、深紫木蓝和檵木的TN出现明显根际效应,但是紫弹树和青檀则没有明显的根际效应(图1)。
2.2 土壤酶活性的季节变化及其根际效应
由表1可见,旱季,6种植物的根际土β-葡聚糖酶(βG)活性和N-乙酰-β-D-葡萄糖苷酶(NAG)活性显著高于非根际土。雨季,除了紫弹树根际土和非根际土的βG酶活性没有显著差异之外,其余树种具有显著差异; 另外,除了檵木的根际土和非根际土NAG酶活性没有显著差异之外,其余树种具有显著差异。
灌木阶段和原生林阶段的根际土和非根际土βG酶和NAG酶活性在雨季大于旱季。
无论是旱季还是雨季,灌木阶段和原生林阶段6种植物的βG酶和NAG酶活性的R/B值(根际土中的值除以非根际土中的值,比值越大则根际效应越明显)均远远大于1。因此,6种植物的两种土壤酶活性具有明显的根际效应。
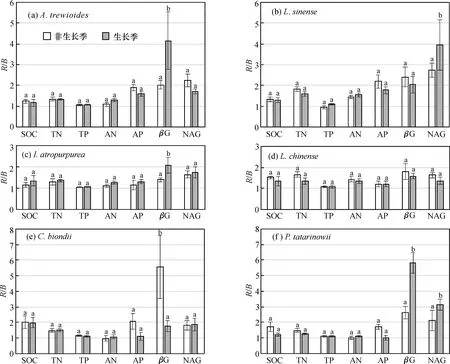
图1 喀斯特地区6种植物土壤养分和酶活性的根际效应(R/B比值)特征Fig.1 Rhizosphere effects (R/B) of soil nutrients and enzymes of six plants in karst region
2.3 土壤养分变化和酶活性的关系
由表2可知,βG酶、NAG酶活性与土壤SOC、TN、TP、AN和AP具有显著的正相关关系;同时,除土壤SOC和AP之间无显著相关关系之外,其余指标之间均有显著的正相关关系。上述结果表明,土壤有机质和氮磷养分总量是氮磷有效性含量变化的基础,而高的土壤酶活性提高了氮磷的有效性含量。由此可见,土壤酶活性的根际效应对植物根际区土壤氮磷养分有效性的提高有积极的促进作用。
3 讨 论
3.1 喀斯特植物根际效应特征
根据第2节研究结果,两个植被恢复阶段6种植物根际土壤的全磷均未出现根际效应,有机碳和全氮的根际效应不明显,有效氮、有效磷、βG和NAG酶活性出现明显的根际效应。植物根际是植物和土壤进行物质和能量交换的重要场所。
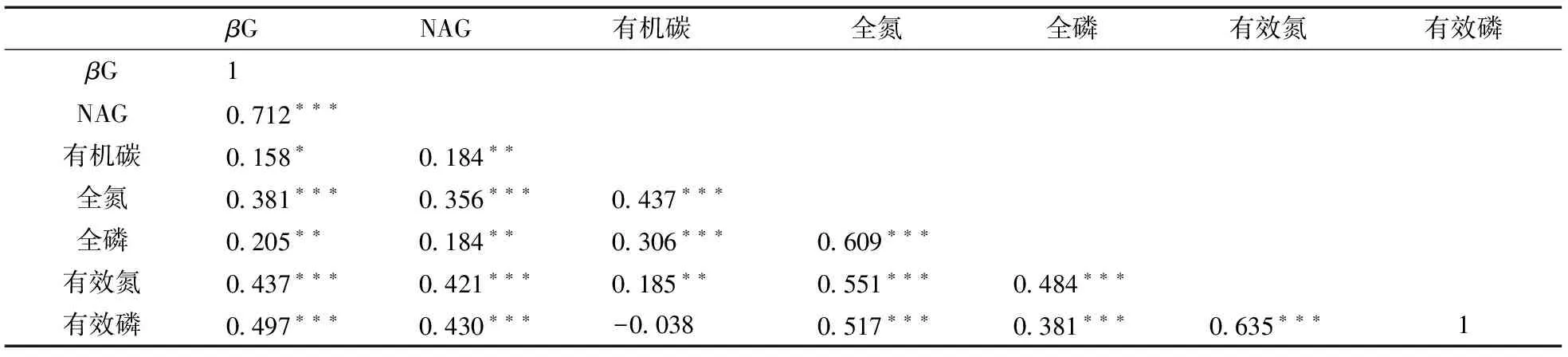
表2 土壤养分含量和有效性与酶活性的相关关系Table 2 Correlation coefficients among soil nutrients and enzymes
注:***,小于0.001;**,小于0.01;*,小于0.05。
一方面,植物地上部分有机物向地下输入时由根系分泌进入根际土壤,这些有机物包括可溶性的糖类、有机酸、氨基酸、酚类和胞外酶等物质[34],一部分供给微生物使用,一部分则会流失掉,还有一部分沉积下来而形成根际效应。根系分泌的可溶性有机物与土壤养分状况具有较强的相关性。一般情况下,当土壤养分丰富时,植物会降低细根分泌物的释放量;而当土壤养分贫瘠时,植物会提高细根分泌物的释放量[35]。当土壤养分含量较低时,植物分泌较多的有机酸和水解酶等有机物,进入土壤后与土壤各种阴阳离子发生氧化还原反应、螯合作用、催化作用等,使土壤有效性养分含量提高[36]。有研究表明,高碳氮含量的有机物分泌显著提高了微生物的呼吸速率和微生物量以及酶活性[37]。另一方面,根系输入土壤的有机物质还包括细根细胞渗出到土壤的低分子量化合物、根毛分泌的高分子粘胶状物质及细根表皮脱落物[3,38]。这些有机物输入土壤后,经过土壤微生物的粉碎和降解,增加了根际区有机碳、氮的沉积,可激活微生物和土壤酶活性,提高氮磷养分的有效性。
综上,根际土有机碳氮、有效性养分和酶活性极易出现根际效应,与本研究得到的结果相一致。但是,根际土全磷在不同植物和不同季节没有出现根际效应(表1)。 通过与多项研究结果进行物种和群落水平的比较(表3),发现喀斯特地区的土壤全磷根际效应值在0.89~1.18,平均值为1.04,表明喀斯特地区的土壤全磷没有出现明显的根际效应。在土壤钙含量较高的喀斯特地区,土壤P容易与Ca结合形成P-Ca化合物,导致该地区土壤磷普遍匮乏[16],可能由此导致该地区不同恢复阶段的植物根际土没有出现全磷的根际效应。但是,植物为了吸收更多的P,可以影响根际区域的生物化学特征,从而使得有效磷的浓度大大高于非根际土,使得根际土的有效磷具有明显的根际效应。因此,喀斯特地区不同恢复阶段的植物根际土没有出现全磷的根际效应,但是有效磷具有根际效应(图1)。
3.2 季节对植物根际效应的影响
季节变化是影响植物根际效应变化的一大因素。从表1可以看出,6种植物在雨季的根际土和非根际土全氮、有效氮、有效磷、βG和NAG酶高于旱季,而全磷在雨季和旱季之间没有显著差异。可能原因为:在雨季,高温多雨的环境有利于植物的生长,导致根系分泌的有机物增加,这种情况在落叶灌木和乔木更加明显,如红背山麻杆、深紫木蓝、檵木、紫弹树和青檀。喀斯特地区具有明显的地质性干旱,在干旱期落叶植物势必会落叶,其各项生理活动必须在雨季高效完成。有研究表明,喀斯特地区落叶植物的光合作用效率和水分利用效率比常绿植物高[43],但是常绿树种的根际土养分含量在雨季和旱季并没有显著差别。尽管旱季的温度、降水量等方面均低于雨季,常绿植物可能更倾向于平稳响应环境变化。
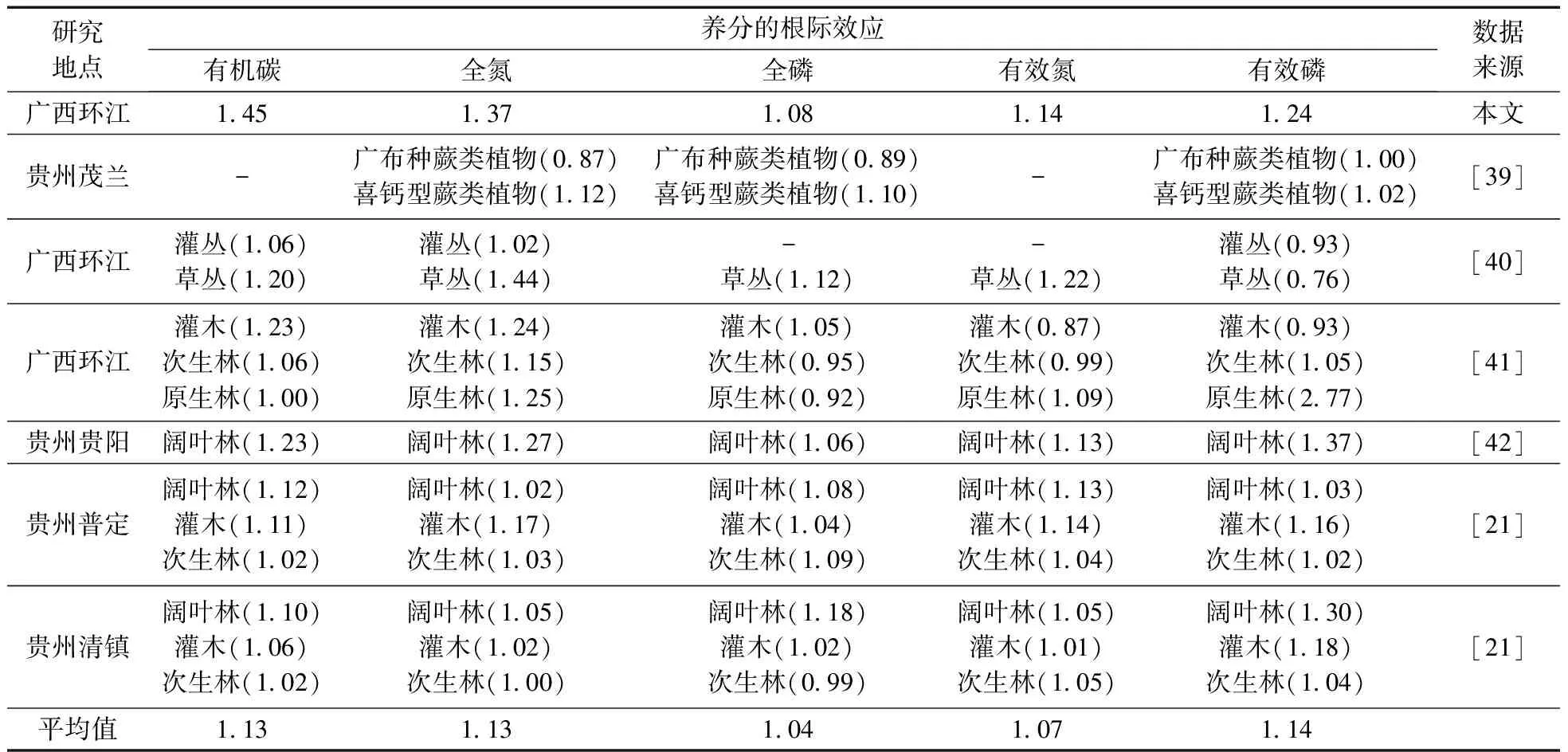
表3 喀斯特地区土壤养分的根际效应(R/B)特征Table 3 Rhizosphere effects (R/B ratios) of soil nutrients in karst ecosystem
上述情况说明,喀斯特地区落叶植物比常绿植物对环境变化的响应要更加迅速,这种响应与植物的光合作用效率、水分利用效率等各种生理活动密切相关。因此,在光合作用、水分利用效率等各种生理活动更高的雨季,喀斯特地区植物的土壤养分和酶活性的根际效应明显高于旱季。
3.3 物种对植物根际效应的影响
植物物种也是影响根际效应变化的另一大因素。原生林阶段乔木(除了檵木)的有机碳、全氮、有效氮、有效磷、βG和NAG酶根际效应高于灌木(表1)。这一现象可能与不同植物的生长速率和养分需求差异有关。在喀斯特地区,植被恢复后期的生物量是恢复前期的10倍左右,植被覆盖度和碳密度大幅增加,养分需求大大提升[12,44-45]。在前期研究中也发现,喀斯特地区植被恢复阶段前期(灌木林)物种受到土壤N和P缺乏的双重限制,而植被恢复阶段后期(原生林)物种受到土壤P缺乏的限制[16]。在土壤养分缺乏的情况下,植物为了能从土壤有机质或者母质中获取更多的养分元素,会诱导并提高根际区土壤的微生物活力和胞外酶活性[31]。βG和NAG酶有利于加快碳氮等元素从土壤有机质中释放,提高土壤中有机碳含量和氮等的有效性[32]。在本研究中也得到类似的结论:土壤βG和NAG酶活性与土壤养分含量和有效性显著正相关(表2)。同时,土壤有机碳含量的提高为微生物生长提供充足的能量,从而提高微生物的数量和活性,进而促进胞外酶的分泌,保证微生物生长和酶分泌两者关系的良好发展。在土壤钙含量较高的喀斯特地区,土壤酶活性(包括磷酸酶、脲酶及其他一些酶)的提高可以使得土壤P从P-Ca化合物中析出,从而提高土壤P有效性。土壤碱性磷酸酶活性随着植被的正向演替而逐步提高[20],而且土壤有机碳和氮含量随着植被正向演替而逐渐提高,土壤肥力逐步改善[26]。因此,植被恢复阶段后期(原生林)乔木的有机碳、全氮、有效氮、有效磷、βG和NAG酶活性根际效应高于植被恢复阶段前期的灌木林树种。另外,大多数物种的βG和NAG酶活性的R/B值高于土壤有效氮和有效磷的根际效应(图1),说明土壤酶活性的根际效应强于土壤有效性养分的根际效应,土壤酶活性是土壤养分有效性提高的重要驱动力之一。
4 结 论
(1)喀斯特生态系统两个植被恢复阶段6种植物的根际土和非根际土的养分有效性含量和酶活性在雨季显著高于旱季,表明喀斯特地区植物根际土养分有效性和酶活性具有明显的季节性变化。
(2)喀斯特地区两个恢复阶段6种植物的AN、AP、βG和NAG酶活性均出现根际效应但TP未出现根际效应且SOC和TN根际效应不明显,表明喀斯特地区植物受土壤氮磷养分限制的情况下,氮磷养分有效性和酶活性的根际效应仍然比较活跃,影响着植物根际区的生物化学特征和养分的吸收效率。
(3)植物根际土壤酶活性与养分全量和有效性含量呈现显著的正相关关系,说明土壤养分全量是养分有效性的基础,而土壤酶活性的根际效应对喀斯特地区植物根际区的氮磷养分有效性提高具有重要作用。
