不同光强对药食两用鳞尾木幼苗生长及光合特性的影响
2020-06-01朱成豪唐健民韦霄高丽梅邹蓉秦惠珍
朱成豪 唐健民 韦霄 高丽梅 邹蓉 秦惠珍
摘要:为探讨适宜鳞尾木这一特有药用野菜生长的光照环境,并为其人工栽培提供有益参考和科学依据,以2年生鳞尾木扦插苗为试验材料,设置10%、30%、50%、100%的光照梯度,研究鳞尾木在不同光强下的生长与光合特性。结果表明:一定程度的遮阴更有利于鳞尾木幼苗的生长,50%光照下鳞尾木的株高、地径及冠幅的增长量均高于其他3种水平;不同光强对鳞尾木的光合特性影响明显,50%光照有利于提高鳞尾木的气孔导度和蒸腾速率。50%遮阴率是鳞尾木生长的适宜条件,而强光或者弱光会对其形成光抑制或弱光胁迫,进而影响其生长。
关键词:鳞尾木;幼苗生长;光合特性;人工栽培;光照
中图分类号: S567.1+90.1文献标志码: A
文章编号:1002-1302(2020)08-0174-05
收稿日期:2019-04-12
基金项目:广西科技基地和人才专项(编号:桂科AD17129022);桂林市科技成果转化与推广项目(编号:20140115-1);广西植物研究所基本业务经费(编号:桂植业18013、桂植业18014)。
作者简介:朱成豪(1994—),男,河南商丘人,硕士研究生,主要从事中药资源开发及检验工作。E-mail:medicalzch@163.com。
通信作者:邹 蓉,副研究员,主要从事保护生物学研究。E-mail:175183030@qq.com。
光照是植物生长发育、形态建成中重要的环境调控因子之一,对植物的生长发育[1]、光合特性[2]、生理代谢[3]、品质形成[4]等具有重要的影响。植物只有在合适的光强下才能更好地生长,光强过弱,植物会出现徒长、叶片变大、变薄等不良症状[5];光强过强,植物则会出现萎蔫、叶片变小、变厚等不良症状[6]。适当改变光照强度可在一定程度上促进植物的生长发育及次生代谢产物的合成。例如:弱光条件有利于喜树碱的合成与积累[7];低光照可提高长春花叶片中文朵灵碱和长春质碱含量,而抑制长春碱的合成[8];75%光照条件可促进喜树植株生长,提高光合速率[9];78%光照条件有利于老鸦瓣产量的积累[10]。植物种类不同对光照强度的需求量及其最佳生长光强也不同,同一植物在不同生长阶段对光强的需求及其最佳光照条件也不同[11]。可见,研究不同光照对植物生长和光合特性的影响对提高植物产量、品质及人工栽培的可实现性尤为重要。
鳞尾木(lepionurus sylvestris Bl.)系山柚子科(Opiliaceae)鳞尾木属(Lepionurus Bl.)灌木或常绿小乔木植物,别称山芥兰、甜菜树[12]。我国有1种,主要分布在云南东南部與广西西南部,多见生长于河谷密林或石缝间隙中,常被当地居民当作一道味道鲜美的野菜食用。鳞尾木不仅可以食用,因其含丰富的营养成分及具有活性的化学成分,也具有极高的药用价值,被用来预防心脑血管疾病、糖尿病、改善肾功能等。目前,关于鳞尾木的研究主要集中于分布地调查研究[13-14]、营养成分研究[15]、抗氧化活性研究[16-17]、核型研究[18]、组织培养研究[19]等。鳞尾木本身资源稀少、人为栽培困难[20],有关光强对鳞尾木的生长动态、光合特性的影响研究尚未见报道。本研究主要探讨不同光照条件下鳞尾木的形态结构与光合特性的差异,以及其生长动态和光合特性对生长环境光强变化的响应和适应机制,进而为鳞尾木的人工栽培提供科学依据。
1 材料与方法
1.1 试验地概况
试验所在地为广西植物研究所特色经济植物研究中心的场地大棚内,110°18′N,25°04′E,海拔175 m,属于亚热带季风气候区。年平均日照时数为1 680 h,年平均气温为23.5 ℃,年平均降水量 1 949.5 mm,年平均无霜期300 d,年平均相对湿度为82%。该地区气候温和,雨量充沛,光照充足。
1.2 试验材料与方法
所用材料为2年生鳞尾木扦插苗,选择幼苗大小基本一致,平均株高17.9 cm。试验于2017年1月20日将鳞尾木移植于10 L塑料花盆中。栽培基质为砂页岩及第四纪红土发育的酸性土壤,pH值4.7~6.5,土壤中氮、钾、镁含量高,有机质含量较高,质地为黏土。植株恢复生长后,于2月20日开始处理,共处理449 d,整个试验期间加强病虫害防治和施肥浇水等管理。试验共进行4个处理,每个处理共10株幼苗。用黑色遮阳网进行遮阴,按照遮阳网的不同密度调节光照强度,建立3层遮阴(L10,约全光照的10%)、2层遮阴(L30,约全光照的30%)、1层遮阴(L50,约全光照的50%)、不遮阴对照组(L100,全光照)4个不同遮阴棚。每个处理10株幼苗,共40株。
1.3 数据采集与处理
1.3.1 数据采集
2018年4月17日至2018年12月17日每隔60 d进行生长检测,每次检测6株,用直尺测量株高和冠幅(东西、南北2个方向),用钢制游标卡尺测量地径,分别记录数据。光合速率的测定选在12月22日08:00,选取不同植株的相同部位的功能叶利用Li-6400(LI-COR,Lincoln,USA)便携式光合作用系统测定。
1.3.2 数据处理
冠幅=东西冠幅×南北冠幅。分别计算6株的地径、株高、冠幅的平均值,按照月份进度对4种不同光照条件进行差异比较。
光响应曲线的测定具体过程如下:测量前将待测叶片在1 000 μmol/(m2·s)光强下诱导30 min(仪器自带的红蓝光源)以充分活化光合系统。使用开放气路,空气流速为0.5 L/min,叶片温度为 28 ℃,CO2浓度为400 μmol/mol(用CO2钢瓶控制浓度)。设定的光照度梯度为2 000、1 800、1 500、1 200、1 000、800、600、400、200、150、100、50、0 μmol/(m2·s),测定时每一光强下停留3 min,以光量子通量密度(PFD)为横轴、净光合速率(Pn)为纵轴绘制光合作用光响应曲线(Pn-PFD曲线),依据Thornley的方法[21]拟合Pn-PFD的曲线方程:
An(I)=αI+Amax-(αI+Amax)2-4θαIAmax2θ-Rd。(1)
式中:An(I)为净光合速率,I为光强,θ为曲线的曲率,α为植物光合作用对光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率,Amax为最大净光合速率,Rd为暗呼吸速率。光补偿点(LCP)和光饱和点(LSP)是利用低光条件下[PFD≤200 μmol/(m2·s)]的Pn与PFD构建线性方程(Pn=mPFD+n,LCP=-n/m,LSP=(Pmax-n)/m)所得[22];表观量子效率(AQY)为0~100 μmol/(m2·s) 光照度范围内净光合速率与光照度直线方程的斜率[23]。
本试验还测定了不同光照处理下的蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)等参数,同时记录光合有效辐射(PAR)和大气CO2浓度(Ca)等环境参数,并据此计算气孔限制值(Ls)=1-Ci/Ca、瞬时水分利用效率(WUE)=Pn/Tr[24],作出显著性差异表进行比较。
借助Excel软件进行原始数据的相关计算,用SPSS 22.0对数据进行one-way ANOVA分析(P<0.05),用光合计算4.1.1软件辅助计算光响应曲线拟合值,利用Sigma-Plot 12.5绘制图形。
2 结果与分析
2.1 不同光照度处理对鳞尾木幼苗生长特性的影响
地径、株高、冠幅是衡量优质苗木出圃的重要标准。由表1可看出,不同光照下,鳞尾木的地径、冠幅存在较大差异(P<0.05)。由图1可知,4种处理以L50处理下生长最佳,地径生长量由最初的0.27 mm生长到12月的0.44 mm,增长了63%,而L100、L30、L10增长了17%、40%、30%,总体表现为L50>L30>L10>L100。各处理的株高生长均呈现不同程度的增长,其中以L50处理下增长趋势最为明显,由4月的12.8 cm生长到了12月的 19.9 cm,增长了55.5%,其次是L30、L10,而L100处理下增长量最低,增长趋势不明显,仅从12.3 cm生长到了17.9 cm,增长了45.5%,总结果为L50>L100>L30>L10。冠幅的生长变化最大,10月到12月的生长趋势降低,可能是由于鳞尾木幼苗落叶或基部老叶脱落,影响了数据的测量导致,整个测量周期内,L100、L50、L30、L10冠幅的增长率分别为8%、33%、25%、15%,结果表现为L50>L30>L10>L100。以上結果说明一定的遮阴会对幼苗生长产生有利影响,本试验中以L50处理对鳞尾木的生长产生最佳影响,可能是由于过分光照会引起植株体内发生光抑制现象,过剩的光能会引起植物体内光化学效率的降低,光照过低又会引起弱光胁迫,抑制花芽分化,从而延缓植物的生长效率。
2.2 不同光照度处理对鳞尾木幼苗光合-光响应曲线的影响
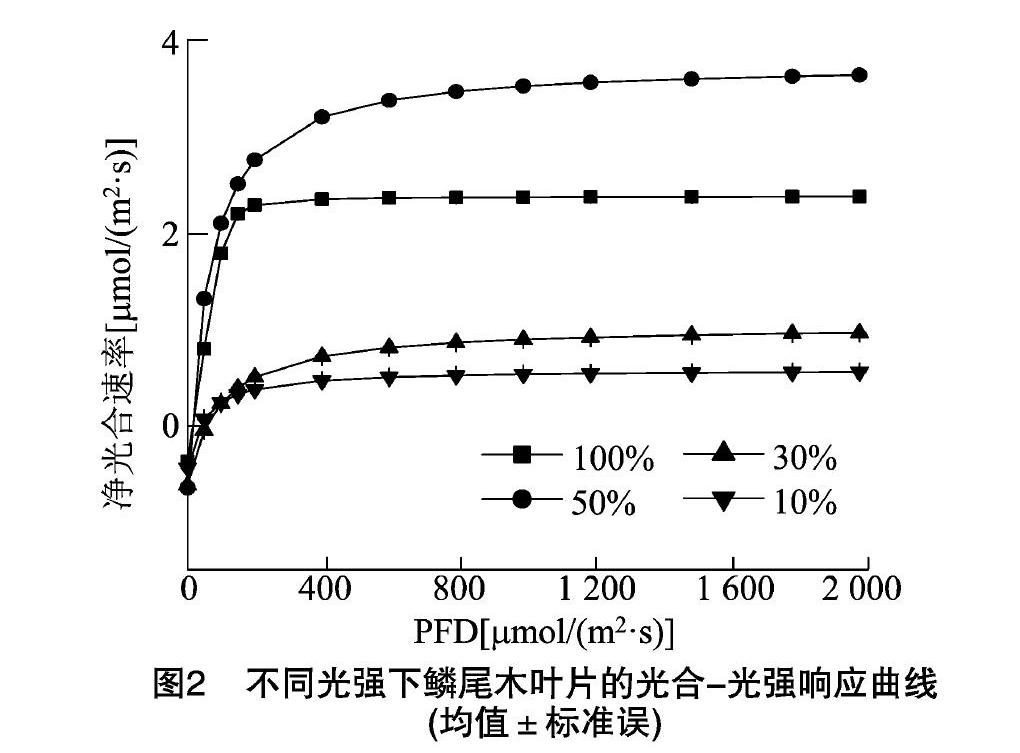
光合作用是反映植物生长好坏的重要指标,而光响应曲线可反映植物Pn随PFD增加的变化规律。由图2可知,不同光照度环境下鳞尾木的光合-光照度响应曲线随光照度的增加变化趋势一致,但Pn大小存在明显差异,其中生长在50%光照下的鳞尾木净光合速率(Pn)最高,其次为100%、30%,10%光照下的净光合速率最低,当PFD为0时,Pn均为负值,随PFD的增加,净光合速率(Pn)逐渐增加,净光合速率低说明同化能力弱,不利于苗木生长,净光合速率高说明同化能力强,能够吸收较多的光能,制造较多的有机物,有利于植株较好地生长。因此50%光照下较适宜鳞尾木的生长。
2.3 不同光照度处理对鳞尾木叶片气体交换参数的影响
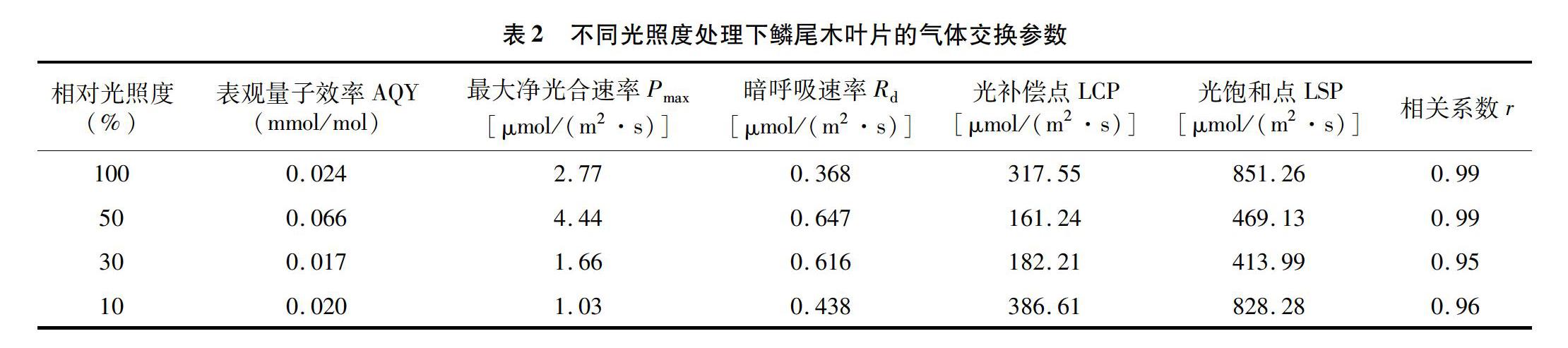
由表2可知,不同处理下鳞尾木LCP差异较大,其中L30和L50处理下的LCP值要低于L10和L100,这说明鳞尾木在前两者环境下达到较小的光照强度就能开始积累干物质,更有利于植株的生长。另外L50处理下生长的鳞尾木最大净光合速率值(Pmax)、暗呼吸速率(Rd)、表观量子效率(AQY)均高于其他处理,这说明随着光照强度的升高,鳞尾木对过强光和过弱光的利用能力都减弱,光照过强或者过弱都不利于其生长。
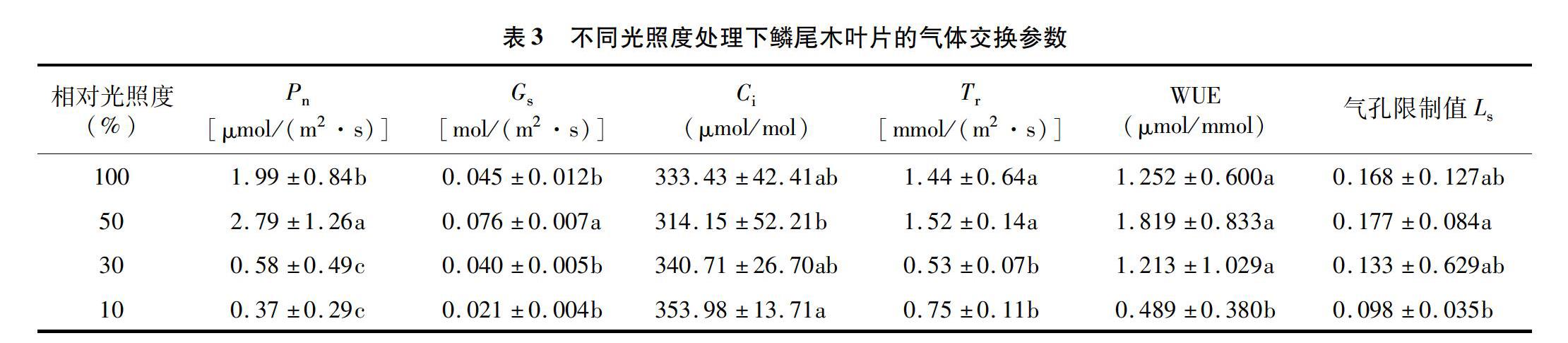
从表3可知,鳞尾木的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、水分利用率(WUE)和气孔限制值(Ls)在不同光照处理下的数据均存在显著差异,除了L50处理下的鳞尾木Ci数据低于其他3种处理外,另5项数据均高于其他3种处理,Ci降低说明叶片固定C的能力提高。植株叶片的气孔导度下降的同时,叶片蒸腾速率也开始下降,而水分利用效率却提高,表明光照的改变,使叶片气孔导度下降,虽然在某种程度上减少了CO2的吸收,影响了光合速率,但气孔导度的下降也伴随水分蒸腾速率的降低,提高保水能力,使水分利用效率得到一定的提高,积累光合产物。
由此得出,鳞尾木在4种不同光照下生长,以50%光照最为适宜。
3 讨论与结论
光照环境会直接影响到植物的代谢分配和光合作用[25],植物长时间内生活在不同光环境下,会以自身变化来适应所处的环境[26],一般来讲,光照增加有利于植物的生长和生物量积累,但一些植物幼苗的生物量积累存在着一个最佳光照度范围,过强的光照反而抑制植物的生长[27],造成叶片灼伤等不利影响,而长期处于过弱的光照环境下,新生出的叶片变小,叶色变浅,植株出现徒长现象。施爱萍等认为50%以上遮阴不利于干簪物质积累,随着光照度的减弱,各生物量(叶、叶柄、根)迅速减少[28-29];对重楼(Paris polyphylla)[30-31]和半夏(Pinellia ternata)[32]的研究也表明其在适宜的遮光下生长良好。本研究表明,不同光照度对鳞尾木幼苗生长的影响明显,50%遮阴处理下,鳞尾木的株高、地径、冠幅均高于其他3种光照处理,这与易伟坚等的研究结果[33-36]基本一致,说明一定程度的遮阴能够促进鳞尾木幼苗的生长,光照过强和过弱均不利于植株生长,可能是由于光照充足引起植株体内产生光抑制现象,严重减缓植株的生长速率[37]。一定遮阴处理下的苗木干物质积累分配机制会根据光照度的变化进而改变植物的生长环境,并影响植物营养物质的吸收及其体内有机产物的重新分配等一系列生理过程,从而更有利于植株各器官的干物质积累[38]。
