亚洲玉米螟幼虫利用反吐液抑制玉米叶片丁布合成的研究
2020-05-23高超男宋鹏飞姚双艳盛子耀李为争罗梅浩原国辉
高超男,宋鹏飞,姚双艳,盛子耀,李为争,罗梅浩,原国辉
(河南农业大学植物保护学院,河南 郑州 450002)
亚洲玉米螟Ostriniafurnacalis(Guenée)是黄淮海流域夏玉米田最严重的蛀穗、蛀茎害虫之一。玉米茎秆被蛀后,营养运输阻断,整体长势衰弱;穗部被蛀后的机械损伤口还易诱发玉米穗腐病等病害[1-2]。一般可以使夏玉米减产20%~30%,严重时减产达30%以上[3]。幼虫的隐蔽取食习性给农药防治带来很大困难[4-5]。因此,探索玉米自身抗虫物质与亚洲玉米螟取食的相互调控机制,对于玉米抗虫良种选育具有重要意义。异羟肟酸类是玉米产生的主要防卫性次生代谢物,典型代表是丁布(2,4-二羟基-7-甲氧基-1,4-苯并恶嗪-3-酮),对植食性昆虫具有广谱抗性[6]。受丁布影响的玉米害虫包括亚洲玉米螟[7-10]、欧洲玉米螟O.nubilalis[11]、西南玉米杆草螟Diatraeagrandiosella[12]、粉红茎螟Sesamianonagrioides[13]、甜菜夜蛾Spodopteraexigua[14]、南方灰翅夜蛾S.eridania[15]、豆秆野螟O.scapulalis和虎杖螟O.latipennis[10]、玉米根萤叶甲Diabroticavirgiferavirgifera[16]等。植物防卫物分为构成性防卫物和诱导性防卫物,前者是植物生长发育过程中自发产生的,后者则是外源因素诱导才会产生的,二者的合成往往是以生物量积累速度延缓为代价的[17]。针对虫害这种偶发性威胁,很显然诱导性防卫对植物而言更有优势[6]。因此,推测禾本科植物中的丁布可能也是虫害诱导产生的化学防卫物。
调控玉米叶片中丁布合成量的外源因子包括物理因子、化学因子、生物因子三大类。物理因子如机械损伤和高温[18]、光照[18-19]、水胁迫[20]等;化学因子如茉莉酸[17]、茉莉酸甲酯[21-22]、水杨酸甲酯[18,22]等;当前生物因子对禾本科作物中丁布含量的影响了解不多,主要包括伴生性杂草[23-24]和昆虫取食[18]。例如,经过大头针机械损伤后,农大108和连玉15的玉米叶片中丁布含量(分别为1.001 567和1.416 224 mg·g-1)和对照(分别为0.905 877和0.948 377 mg·g-1)相比均显著增加;但2个品种3龄亚洲玉米螟幼虫取食损伤的反应却不同,虫害损伤的农大108叶片丁布含量(1.282 654 mg·g-1)极显著高于对照,但连玉15经亚洲玉米螟取食损伤后丁布含量(1.024 665 mg·g-1)和对照相比却没有显著差异。丁布的主要生态学功能是防卫植食性昆虫。然而,植食性昆虫这种潜在生物因子对丁布生物合成的逆向调控作用目前尚缺乏系统的研究,尤其是丁布合成量与害虫种群密度、取食持续时间、叶片损伤程度等因素之间的关系。本研究通过机械损伤、亚洲玉米螟取食和模拟取食等诱导处理玉米叶片,每个处理内部各设置不同的诱导时间,然后测定玉米叶片丁布含量的变化,以便弄清丁布是虫害诱导性还是构成性化学防卫物质,为抗虫玉米品种的选育提供科学依据。
1 材料与方法
1.1 供试昆虫
亚洲玉米螟为河南农业大学昆虫生态实验室饲养(14 L∶10 D,(27±1)℃,相对湿度65%~85% )。饲料制备方法为:取716.2 g蒸馏水,加入11.5 g琼脂、2.9 g山梨酸后煮沸,然后加入86 g玉米粉、86 g大豆粉、51.6 g酵母粉和43 g葡萄糖,充分搅拌,物料降低至40℃再加入2.9 g 维生素C,并充分搅拌后降低至室温备用[25]。羽化后成虫饲喂浸渍于脱脂棉球上的10%蔗糖溶液,用载玻片作为产卵基质。
1.2 玉米品种
供试品种为“浚单20”,购于河南秋乐种业有限公司。播种于田间防虫网内的花盆中,每盆种植1株,以避免其他害虫取食以及不同玉米植株根系之间的相互作用对丁布合成量的潜在影响。大喇叭口期时,将被试植株连同花盆转移到隔离的小网室中,选取苗高和叶片面积大致一致的植株,进行外源因素的诱导处理。
1.3 供试试剂
磷酸、乙醚、甲醇、无水乙酸购买于天津科密欧公司,色谱纯,纯度均在99.5%以上。丁布标样在河南农业大学应用昆虫生态学实验室提取和分离,并在郑州大学分析测试中心采用Agilent 1200/6220高效液相色谱-质谱联用仪检验纯度。
1.4 玉米叶片诱导
为了探讨亚洲玉米螟取食玉米叶时物理因子和化学因子的相对影响,设置下述3组处理。(1)昆虫取食组。取人工饲料饲养的亚洲玉米螟4龄幼虫,用软毛刷接于玉米心叶处。分为1、2、3头·株-13个水平;每种取食处理的诱导时间均分5个水平,即诱导3、6、12、24、48 h。昆虫释放之后用透明塑料圆筒罩住每盆玉米,一是防止幼虫逃逸,二是防止不同植株之间的气味通讯,三是尽量使生长环境接近自然。(2)机械损伤组。亚洲玉米螟为害造成“排孔”症状,故用0.4 cm ID打孔器对玉米苗上部第2片叶做打孔处理,模拟为害所致机械损伤。试验设计基本同上,差别仅在于用叶片打孔的方式代替释放幼虫。打孔数也分为3个水平,即在叶脉两侧分别打4、6、8个圆孔(圆孔直径为0.4 cm)。其中,叶脉每侧打4个圆孔的处理,圆孔均在一排上;叶脉每侧打6个圆孔和8个圆孔的处理,圆孔分为2排,间距大约4 cm。预试验表明,这些圆孔所致损伤叶面积分别相当于幼虫在1、2、3头·株-1释放密度下48 h取食的叶面积。(3)模拟取食组。该组的目的是将取食物理因子(蛀孔)和化学因子(口器反吐液)叠加,并与上述2组丁布含量指标对比。幼虫反吐液收集方法是,在体视解剖镜下用镊子轻压5龄幼虫胸部背面,用1.0 mm内径的毛细管收集反吐液,立刻吹入1.5 mL离心管,多次收集合并的反吐液置于-20 ℃冰箱中保存备用。试验设计基本同上述2组,差别在于叶片打孔后,每个孔周围用移液器均匀涂抹幼虫反吐液2.0 μL。另外,在相同生态环境中各设置1组对照植株,用于化学分析采样的对照植株与不同取食诱导时间相对应。
1.5 丁布提取和分析
对上述不同诱导处理的玉米叶片进行丁布含量分析。为了避免离体叶片自发性代谢变化,野外采集叶片之后,迅速置于独立的冰盒中携带到室内,马上进行丁布提取和色谱分析。
丁布提取方法在WOODWARD等[26]方法的基础上改进,主要改进点是用磷酸缓冲液调节pH值以便提高丁布得率。首先,以亚洲玉米螟取食痕迹或打孔器所致损伤位置为中心,取玉米叶片样品1 g,加少许蒸馏水研磨匀浆,室温静置15 min。然后,在Hanna Hi98128笔式酸度计的实时监测下,用0.1 mol·L-1磷酸溶液将pH值调节至3.0,转移至5.0 mL的离心管中,在12 000 r·min-1转速下离心15 min。最后用移液器取出上清液,每次用等体积乙醚萃取,共萃取3次,在Heidolph-4011型旋转蒸发仪(热源温度40 ℃)上蒸干乙醚,溶于甲醇并过0.45 μm滤膜,在-20 ℃下保存备用。
采用配置C18柱(150 mm × 4.6 mm)的Agilent 1200/6220高效液相色谱-质谱联用仪测定丁布含量。进样量10 μL,柱温28 ℃,流动相为体积比为1∶1的甲醇与0.5%乙酸溶液,柱流速为1.0 mL·min-1,检测波长为262 nm。将色谱峰保留时间、紫外光谱与丁布标样对比。外标法定量,以不同浓度丁布标样的吸收峰面积作出标准曲线,计算不同样品中丁布含量,以每克鲜组织中含有的丁布表示(μg·g-1)。诱导处理类型×损伤强度(打孔数或昆虫释放头数)×诱导时间的每个组合重复3次,各组配置的对照也重复3次,共144份样品。
1.6 数据分析
采用SPSS 19.0 for Windows的一般线性模型(General linear model)统计。首先,采用三因素方差分析比较不同类型的外源因素诱导因子对玉米叶中丁布含量的影响。由于昆虫取食组玉米叶片被食叶面积随诱导时间延长逐渐增大,而机械损伤组和模拟取食组叶损伤面积是定值。因此,统一以有序变量“损伤强度”表示,分为“轻度受损”“中度受损”和“严重受损”3个等级,和相应幼虫释放头数和打孔数对应。该模型自变量为外源诱导因子类型(名义变量)、诱导时间(连续变量)和损伤强度(有序变量),考察所有因素主效应和二阶交互效应。此外,通过3组二因素方差分析更详细地探明昆虫取食组、机械损伤组和模拟取食组各组内部不同处理条件的具体影响,昆虫取食组的自变量为试虫释放头数和诱导时间,其他2组的自变量为打孔数和诱导时间。多重比较采用Student-Newman-Keuls法,显著性水平取α=0.05。
2 结果与分析
2.1 不同外源诱导因素对玉米叶片丁布含量的影响
外源诱导因子类型对玉米叶片中丁布含量有极显著影响(F2,114=31.87,P<0.000 1),其他2个主效应的影响也达到显著水平(诱导时间:F4,114=3.03,P=0.020 5;损伤强度:F2,114=4.55,P=0.012 6),诱导因子类型和诱导时间之间存在极显著交互(F8,114=5.77,P<0.000 1)。损伤强度和另外两个因素之间则均不存在显著交互作用(损伤强度×诱导因子类型:F4,114=2.15,P=0.079 2;损伤强度×诱导时间:F8,114=0.91,P=0.514 4)。
3种因素主效应的多重比较如表1所示。诱导因子类型比较中,机械损伤组玉米叶片中丁布含量最高为(492.39±53.00)μg·g-1,且显著高于其他3组;对照组丁布含量为最低(419.55±30.53)μg·g-1,且显著低于其他3组;昆虫取食组和模拟取食组的玉米叶片含量居中且无显著性差异。受损伤玉米叶片丁布含量均显著高于健康玉米叶片,但与受损程度无关;类似地,各诱导处理时间的玉米叶片丁布含量均显著高于对照,但凡是经过外源因素诱导之后的叶片,丁布含量均与诱导时长无关。
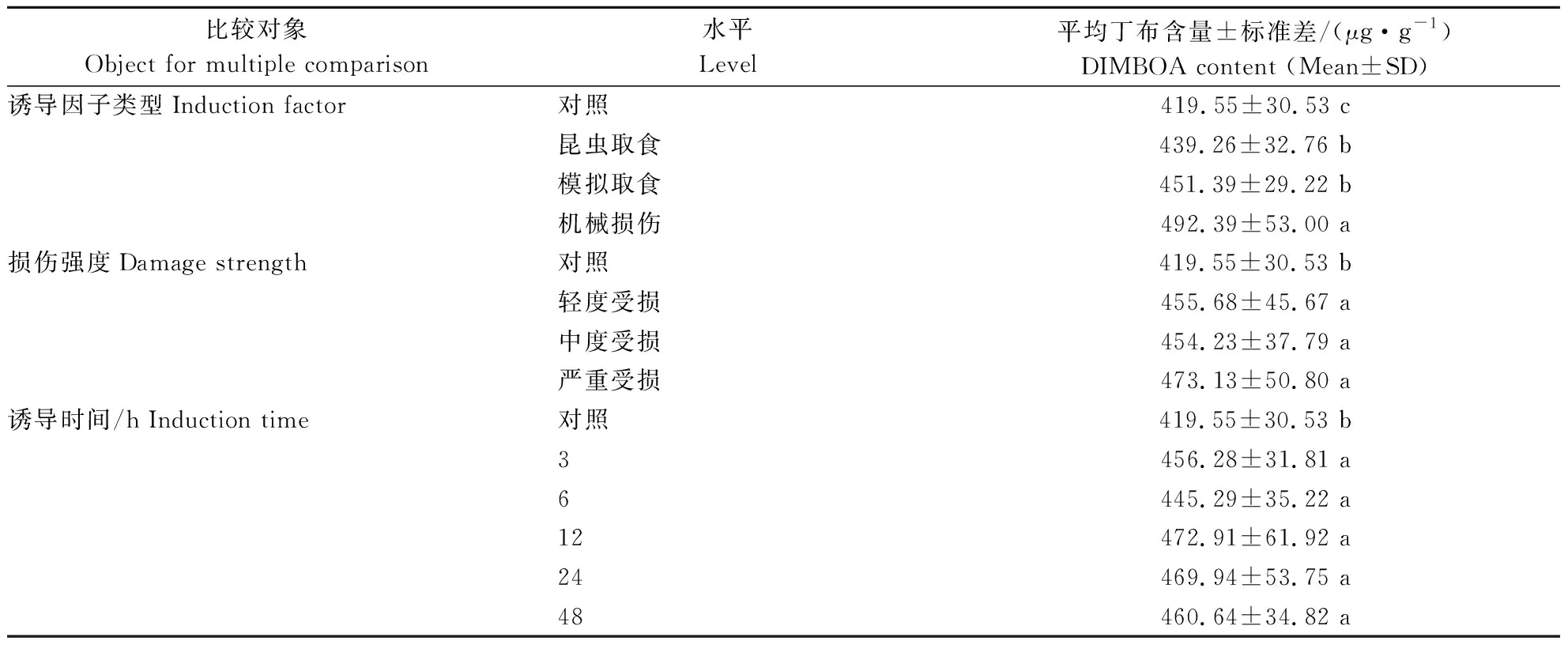
表1 外源诱导因素的不同水平对玉米叶中丁布含量的影响Table 1 Effect of different levels of exogenous induction factors on DIMBOA content in maize leaves
注:平均丁布含量一列中,不同小写字母表示相同比较对象内各水平之间的差异显著(P<0.05)。
Note: In the column of mean DIMBOA content,the lowercase letters attached to the numbers indicate significant difference between levels within the same object for multiple comparison(P<0.05).
2.2 亚洲玉米螟取食对玉米叶片丁布含量的影响
亚洲玉米螟释放头数(F2,32=0.41,P=0.670 3)和取食诱导时间(F4,32=2.48,P=0.063 8)对玉米叶片丁布含量均无显著影响,二者也不存在显著交互(F8,32=0.08,P=0.999 5),无需多重比较。
2.3 机械损伤对玉米叶片丁布含量的影响
机械损伤组中,诱导时间对玉米叶片中丁布含量有极显著影响(F(4,32)=7.96,P=0.000 1),打孔损伤程度的影响也达到了显著水平(F(4,32)=5.09,P=0.012 0),但二者不存在显著交互(F(4,32)=0.66,P=0.723 4)。多重比较表明,打孔数效应的显著性主要表现在叶片打孔与非打孔方面,而不是打孔数量(表2)。诱导时间是机械损伤组中影响玉米叶中丁布含量的主要因素,机械损伤后6 h内丁布含量差异不显著,丁布合成高峰在机械损伤诱导12 h后逐渐下降(表2)。

表2 机械损伤后不同诱导时间和损伤强度对玉米叶中丁布含量影响的多重比较Table 2 SNK multiple range test of DIMBOA content in maize leaves exposed to mechanical damage
注:本表为诱导时间和打孔数(损伤强度)两个主效应的多重比较结果。分析诱导时间时,相同时间点不同打孔数诱导的丁布含量数据合并分析;相应地,分析损伤强度时,相同打孔数不同诱导时间的丁布含量数据合并分析。取样到上机分析的时滞最多1 h。平均丁布含量一列中,数字之后的小写字母表示相同比较对象内各水平之间的差异显著性。
Note: This table indicates the multiple-range comparison of two main factors (Induction time and Hole number).When the effect of induction time was analyzed,the DIMBOA content data from maize leaves induced with different hole numbers at the same time point were merged for analysis; correspondingly,when the effect of hole number was analyzed,the DIMBOA content data from maize leaves induced with the same hole number at different times were merged for analysis.The time lag from sample collection to chemical analysis is not longer than 1 h.In the column of mean DIMBOA content,the lowercase letters attached to the numbers indicate significant difference between levels within the object for multiple comparison.
2.4 模拟取食对玉米叶片丁布含量的影响
在机械打孔处添加亚洲玉米螟反吐液后,丁布含量表现出和正常昆虫取食组相似的模式,诱导时间(F(4,32)=0.55,P=0.699 9)、打孔数(F(2,32)=1.10,P=0.344 5)及二者交互作用(F(8,32)=0.51,P=0.842 7)均不显著,无需多重比较。
2.5 不同外源诱导因素对玉米叶片丁布含量影响的时相分析
从图1可以看出,玉米叶片对不同外源因子诱导的短期反应(0~6 h),均是小幅增加了丁布合成量。然而,随着诱导时间延长,玉米对3类外源诱导因子的响应模式明显不同。机械损伤造成丁布含量大幅上升,12 h时达到顶峰,随后缓慢衰减,一直到试验设计的最大诱导时间(48 h)时仍比其他2组丁布含量高。而昆虫取食组和模拟取食组响应模式相似,被诱导玉米叶片丁布含量从玉米短期反应期(3 h)过后,一直维持在和对照无显著差异的水平。不同点在于,昆虫取食组丁布含量缓慢上升,而模拟取食组丁布含量缓慢下降。

图1 不同外源因素类型与诱导时间的交互作用分析Fig.1 Interaction analysis of different types of exogenous induction factors and induction times
3 结论与讨论
大田玉米品种丁布含量与抗螟性关系的数据,不能确定是成虫寄主选择过程的差异所致,还是幼虫取食造成特定植株丁布含量变化所致[9],而另一些研究是将丁布或结构类似物定量混入人工饲料进行测试[3,8,10,12,15],难以弄清玉米和亚洲玉米螟之间的适应和反适应对策。通过研究机械损伤、昆虫取食和模拟取食对玉米叶片丁布含量的影响,表明诱导因素类型占主导作用,诱导时间和损伤强度从属于前者发挥作用。研究结果表明,只有机械损伤刺激丁布大量合成,诱导后12 h达到高峰,然后逐渐下降。亚洲玉米螟取食并不会诱导丁布合成量大幅增加,相反,会起到显著抑制作用。
亚洲玉米螟幼虫所蛀孔径随着持续取食逐渐扩大,尽管打孔器孔径0.4 cm尽量模拟“排孔”,仍不能排除“一次性损伤”和“逐渐损伤”造成玉米应激反应速度的差异,单纯比较昆虫取食组和机械损伤组也许会得出这样的结论。然而,模拟取食组是用相同打孔器处理的,不同点仅在于每孔周围涂布了20 μL幼虫反吐液,就造成该组和机械损伤组丁布生物合成模式的显著差异,而和昆虫取食组丁布合成模式相似。因此,可以认为是亚洲玉米螟幼虫反吐液抑制了丁布合成。由于昆虫取食组和模拟取食组内部也设置了对照植株,但昆虫释放头数、打孔数和诱导时间等都不影响丁布含量,故幼虫反吐液能够将丁布合成量维持在“正常”水平上。另外,昆虫取食组丁布含量缓慢上升,而模拟取食组丁布含量缓慢下降,可能是由于实际取食存在着玉米叶受损面积逐渐增加的过程。因此,丁布并非亚洲玉米螟幼虫为害诱导产生的玉米防卫物,而是玉米在各种胁迫下自发合成的[19-20]。健康幼苗不能检测到游离的丁布,只存在稳定的β-葡萄糖苷键合物[27],本研究发现机械损伤是玉米苗丁布合成的主要诱导因素,这与CAMBIER等[27]的结论一致。然而,也有报道玉米受到亚洲玉米螟[18]、西南玉米杆草螟[28]、欧洲玉米螟[29]危害后丁布含量显著增加,可能是由于没有将昆虫取食按照诱导时间系统展开研究造成的。
自然界中昆虫面临着大量时空异质性分布的植物防卫物,对植物化合物抗性的程度和本质随着植物基因型的地理分布、植物-昆虫关系的特异性、局部生态成分而变化。昆虫适应植物防卫物的机制包括嗅觉和味觉回避、排泄、储存、解毒和靶标突变等[30]。玉米鳞翅目害虫适应机制已有报道。玉米心叶中的刺激取食剂会掩蔽丁布对亚洲玉米螟的拒食效果[8],类似报道是南方灰翅夜蛾[14]。亚洲玉米螟[6]和东方粘虫Mythimnaseparate(Walker)[31]消化道中存在的尿苷二磷酸葡萄糖转移酶或其他尿苷二磷酸葡萄糖依赖性酶可以快速将丁布降解为无毒产物,而欧洲玉米螟可以通过排泄的调控将组织中的丁布浓度和虫粪中的浓度比稳定在0.25左右[32]。
鳞翅目幼虫反吐液的功能也已有报道。鳞翅目幼虫取食植物伴随的反吐液影响受害植物与后继植食者之间的相互作用[33]。甜菜夜蛾幼虫反吐液中分离出一种被称为“Volicitin”的物质也能够吸引天敌[34]。然而,鳞翅目幼虫反吐液对植物非挥发性次生防卫物合成的影响少有报道,本研究探索了鳞翅目幼虫利用反吐液从植物防卫物生物合成源头上适应寄主的新机制。
