Variations in fine root dynamics and turnover rates in five forest types in northeastern China
2020-05-19NanWangChuankuanWangXiankuiQuan
Nan Wang · Chuankuan Wang · Xiankui Quan
Abstract Quantifying fine root (≤ 2.0 mm in diameter) distribution and turnover is essential for accurately estimating forest carbon budgets. However, fine root dynamics are poorly understood, possibly because of their inaccessibility. This study quantifi es fine root distribution and turnover rates for five representative Chinese temperate forests types. Fine root number, diameter, biomass, necromass, production, mortality, and turnover rates were measured using a minirhizotron over a 12-month period. More than 90% of the fine roots were < 0.5 mm in diameter, with thin fine roots at shallow layers, and thicker ones in deeper soil layers. The fine root dynamics were significantly different among the forest types. Coniferous plantations had fewer fine roots, less biomass, necromass, production and mortality but greater average diameters than fine roots of broadleaved forests. All traits, except for diameter, decreased along the soil profile. Fine root numbers and production exhibited a unimodal seasonal pattern with peaks occurring in summer, whereas biomass, necromass and mortality progressively increased over the growing season. The turnover rates of roots < 0.5 mm varied from 0.4 to 1.0 a -1 for the five forest types, 0.5-1.0 a -1 for the soil layers and 0.2-1.1 a -1 for the seasons, with the largest turnover rate at the 0-10 cm depth in summer. The patterns of fine root numbers, biomass, necromass, production, mortality, and turnover rates varied with forest types, soil depths, growing season and diameter classes. This study highlights the importance of forest types and diameters in quantifying fine root turnover rates.
Keywords Biomass · Necromass · Mortality · Production · Root dia meter · Root numbe r
Introduction
Fine roots (≤ 2 mm in diameter), functioning for water absorption and nutrient capture, make up one-third of the global net primary production (Jackson et al. 1997; Du and Wei 2018), and are sensitive to environmental changes (Gill and Jackson 2000). Quantifying fine root characteristics and rates of turnover is essential for accurately estimating forest carbon budgets and improving our understanding of belowground processes and their adaption to environmental changes (Eissenstat and Yanai 1997; Clemmensen et al. 2013; Wang et al. 2017). However, fine root dynamics are a least understood aspect of forest ecology, possibly because of their relative inaccessibility. The temperate forests in northeastern China account for one-third of the forest area and standing stock in the country and play a crucial role in national carbon budgets (Wang 2006). The belowground carbon pool and fl uxes in these forests, however, have not been quantified (Wang et al. 2006a; Zhang and Wang 2010).
The first objective of this study was to compare fine root characteristics among five forest types, soil depths and seasons. It has been reported that root characteristics vary with tree species (Kong et al. 2014; McCormack et al. 2015). For example, the fine roots of coniferous species tend to have greater diameters than those of broadleaved species, and those penetrating deeper soil layers are likely to be thicker (López et al. 2001; Pregitzer et al. 2002; Wang et al. 2006b; Kong et al. 2014). Fine root features in the forest levels, however, may differ for each species because plants in natural ecosystems have evolved morphological and physiological attributes in response to resource competition from other plants (Brassard et al. 2009). To maximize exploitation of belowground resources, coexisting species often develop compensating functional traits to partially or completely segregate their niches (Man and Lieff ers 1999; Bennett et al. 2002). Brassard et al. ( 2009) hypothesized that mixed species stands with different functional root traits may attain greater root biomass and production through more effi cient and complete usage of soil resources than a monoculture or species mixture with roots with similar functional traits. However, this hypothesis requires testing. Additionally, fine roots demonstrate plasticity with changes to the environment (Eissenstat and Yanai 1997; Eissenstat et al. 2000; Gill and Jackson 2000), which further complicates our understanding of fine root characteristics. Fine root development is infl uenced by growing season and soil depth (Hendrick and Pregitzer 1992; Satomura et al. 2006; Espeleta et al. 2009; Kou et al. 2015), however, growth and infl uencing factors remain unclear. McCormack et al. ( 2015) noted that the traditional fine-root pool (≤ 2 mm in diameter), is a heterogeneous group of roots which had obvious functional divergence between root diameter classes. In this study, the dynamics and production between root diameter classes are examined. It is hypothesized that the two coniferous plantations will have greater fine root diameters but less biomass, necromass (dead organic material), production, and mortality than the three natural broadleaved stands, and that roots (≤ 0.5 mm) in the surface soil layers will be more numerous in the summer and have greater biomass, necromass, production and mortality than fine roots in deeper soil layers, in other seasons but have larger diameters (0.5-2 mm).
The second objective was to quantify fine root turnover rate (TR) and its spatiotemporal pattern for the five forest types. Fine root turnover is a major pathway for carbon and nutrient cycling in forest ecosystems. It varies with tree species (Steele et al. 1997), growing season (Pregitzer et al. 1993), temperature (Pregitzer et al. 2000), soil depth (Fahey and Hughes 1994; Wang et al. 2017), and fine root diameter (Wells and Eissenstat 2001). Fine root turnover or longevity is commonly estimated with a minirhizotron imaging system in which the TR is calculated from the fine root length by assuming that all fine roots have a uniform diameter or density. However, fine root diameters and densities change with species (Pregitzer et al. 2002; Wang et al. 2006b), seasons and soil depths (Eissenstat and Yanai 1997; Zhang et al. 2009) and function (McCormack et al. 2015). These factors may introduce bias in estimating fine root turnover rates in the minirhizotron approach. For example, when separating fine roots (≤ 2 mm) into absorptive fine roots (≤ 0.5 mm) and transport fine roots (0.5-2 mm), McCormack et al. ( 2015) estimated that fine-root production and turnover accounted for approximately 22% of the global terrestrial net primary production-an approximate 30% reduction from previous estimates, assuming all roots ≤ 2 mm in diameter had the same turnover rate (1.0 a -1 ). It was hypothesized that turnover rates will differ with forest type, at different soil depths and in different seasons because fine root diameters and densities change.
Materials and methods
This study was conducted at the Maoershan Forest Ecosystem Research Station, northeastern China (45° 24′ N, 127° 40′ E). Average altitude is 400 m a.s.l. average slope is 10°-15°. The parent soil material is granite bedrock and the soil is a dark brown medium loam. The climate is continental monsoon and based on meteorological data from 1989 to 2009, the mean annual precipitation is 629 mm and the average annual evaporation is 864 mm. Average annual, January, and July temperatures are 3.1 °C, - 18.5 °C, and 22.0 °C, respectively. The frost-free period is between 120 and 140 days with early frosts in September and late frosts in May. The stands are naturally regenerated or have been planted following large-scale industrial logging of an original primary temperate mixed forest.
The experimental design included five forest types with three random replicated 20 m × 30 m plots in each type. The forest types, which are representative of this region, are: a Korean pine ( Pinus koraiensis Sieb. et Zucc.) plantation, a Dahurian larch ( Larix gmelinii Rupr.) plantation, a Mongolian oak ( Quercus mongolica Fisch.) forest, an aspen-birch ( Populous davidiana Dode and Betula platyphylla Suk.) forest, and a hardwood forest dominated by Fraxinus mandshurica Rupr., Juglans mandshurica Maxim., and Phellodendron amurense Rupr. (Table 1).
Minirhizotron measurements of fine root demography
Dynamics of fine roots were monitored with the minirhizotron research (MR) method (Johnson et al. 2001). Five acrylic MR tubes (90 cm × 5.1 cm inner diameter) were installed randomly in each plot in the autumn of 2006. They were partly inserted at a 45° angle off , reaching a 45-cm depth. Each tube had 45 (1.8 cm × 1.4 cm) rectangular frames sequentially numbered along its length from the bottom to the top. One year after the MR tubes were installed, as recommended by Johnson et al. ( 2001), fine root images were collected from each tube. In this study, the images for analysis were taken with a micro-video camera (Bartz Technology, Santa Barbara, CA, USA) every two weeks from October 15th 2008 to October 15th 2009, a total of 14 sampling times. Images were not taken in winter months (November 16th 2008 to March 14th 2009) because of a thick snowpack (~ 40 cm) and frozen soils. During each sampling time, 45 MR images were taken from the top to the bottom of each tube.

Table 1 Site conditions and overstory characteristics for the five forest types
The images were digitized using the Rootracker software (version 2.0, Duke University Phytotron, Durham, NC, USA). Morphology (length, diameter, volume) and category (living: brown and elastic; dead: black and inelastic; lost: dead or eaten by insects) were recorded. The soil profile was divided intofour layers: 0-10 cm, 10-20 cm, 20-30 cm, and 30-45 cm. For each MR tube, the length and volume of fine roots for each root class and soil layer were averaged. Production and mortality between the sampling times were calculated by summing up all new root lengths per volume and dead root lengths per volume for each MR tube at the end of the period. Lost roots which had been present in the previous measurement were included as a part of mortality.
Measurements of Specific root length
Eight Soilcore samples (10-cm inner diameter) were randomly taken in each plot in June, August, and September of 2009, corresponding approximately to leaf emergence, maximum leaf area, and leaf senescence, respectively. The Soilcores were removed at 0-10 cm, 10-20 cm, 20-30 cm, and 30-45 cm, placed in a cooler (~ 4 °C), transported to the laboratory and kept frozen before they were analyzed.
Fine roots were removed manually from the Soilcores according to depth and washed and sorted into living or dead roots into < 0.5 mm, 0.5-1.0 mm, and 1.0-2.0 mm diameter class. For each class, the roots length were measured with a digital vernier caliper, placed in labeled envelopes, ovendried to a constant mass at 65 °C, and weighed to the nearest 0.0001 g. For each diameter class by soil depth for each forest type, the Specific root length (SRL) was calculated as root length divided by root dry mass (m/g).
Measurements of s oil pr operties
Continuous soil temperature (T5) and water content (W5) at the 5-cm depths in the oak,hardwood, and pine forests were measured with a CR23X datalogger (Campbell Scientifi c Ltd., Logan, UT, USA) attached to thermocouples and water content refl ectometers (CS616-L, Campbell Scientifi c). The data were recorded at 15 min intervals. T5and W5in the larch and aspen-birch forests were derived from the continuous measurements following the protocol of Wang et al. ( 2006a).
To determine Soilchemical properties, five Soilcores in each plot were randomly sampled every month from May to November 2009, and the cores were divided intofour depths, 0-10 cm, 10-20 cm, 20-30 cm, and 30-45 cm. For each depth, total nitrogen (N), phosphorus (P) and potassium (K) were measured only in August; ammonium (NH4+), nitrate (NO3- ), available phosphorus (PA) and available potassium (KA) were measured monthly from May to November. The soil samples were thoroughly mixed, sieved (< 2 mm) and the subsample extracted with 2 M KCl for determining initial NH4+and NO3-concentrations. After homogenization of the air-dried samples in a ball mill, total N was determined by the Kjeldhal method using a Kjeltec 8400 analyzer (Foss Tecator AB, Höganäs Sweden); total P was determined by the sodium hydroxide-molybdenum stibium anti-color method; total K was determined by the sodium hydroxide-fl ame photometry method. Available K levels were determined using ammonium acetate-fl ame photometry and available P concentration by the acid melt-molybdenum stibium anti-color method.
Data anal ysis
Based on vegetation phenology, the growing season was divided into three periods: spring (March-May) when the leaves were out, summer (June to early August), and autumn (mid-August to October) when the leaves commenced falling. The experimental unit was the plot, and the means and standard errors of fine root traits were calculated by averaging plot-level measurements. Fine root turnover rate (TR a-1) was calculated as:where P, M, and B are fine root production (g a -1 ), mortality (g a -1 ), and mean annual standing biomass (g), respectively. Biomass-based production and mortality were calculated as root length divided by Specific root length (SRL).

The analysis of variance with the least signifi cant difference (LSD) test was applied to determine signifi cant Differences in fine root diameter, number, biomass, necromass, production, mortality, Specific root length (SRL), and turnover rates (TR) among forest types, soil depths, and seasons. A correlation analysis was applied to test the relationships between fine root traits and soil properties. All data analyses used the SPSS version 13.0 (SPSS Institute, Chicago, IL, USA).
Results
Specific root length (SRL)
SRL was significantly different ( P < 0.05) among the five forest types for all three diameter classes (Tables 2, 3). The SRL ranking varied with the classes; for the < 0.5 mm class, mean SRL varied from 22.5 m g -1 for the larch forest to 40.5 m g -1 for the pine forest; for the 0.5-1.0 mm class, the average SRL ranged from 14.5 m g -1 for the oak forest to 31.3 m g -1 for the pine forest. The average Specific root length for the 1.0-2.0 cm class ranged from 1.7 m g-1for the pine forest and 4.0 m g -1 for the aspen-birch forest.

Table 2 Three-way analysis of variance for Specific root length (SRL) of different diameter classes

Table 3 Specific root length (SRL) of fine root by forest types, soil depth and season
SRL for the < 0.5 mm class was significantly lower in the 30-45 cm depth than for other depths, varying from 26.6 to 35.6 m g -1 (Table 3). There were signifi cant differences in the SRL for the other two diameter classes among soil depths (Tables 2, 3). The average SRL varied from 17.5-25.2 m g-1and 2.7-3.5 m g-1for the 0.5-1.0 mm and 1.0-2.0 mm classes, respectively. Specific root lengths were significantly different among the seasons for all three diameter classes (Tables 2, 3). Fine roots developed in August and September had lower lengths than those in June. The mean SRL varied from 27.9-52.8 m g -1 , 8.5-25.6 m g -1 and 1.8-3.0 m g -1 for the < 0.5 mm, 0.5-1.0 mm and 1.0-2.0 mm classes, respectively (Table 3).
Fine root diameter and number
Fine root diameter showed a highly right-skewed distribution for all forests, soil depths, and seasons (Fig. 1). More than 90% of the diameters were < 0.5 mm, and 98% < 1 mm. However, the peak frequency of the distribution and the diameter were substantially different among the stands (Fig. 1 A), whereas they were more uniform with soil depth and seasons (Fig. 1 B, C). Peak frequencies of root diameter and distribution in the oak and aspen-birch forests were greater than in the pine and larch forests, whereas peak diameters of the former were less than in the pine and larch forests (Fig. 1 A). In spite of a consistent right-skewed diameter distribution along the soil profile, relatively thicker fine roots (1-2 mm) were found at the 30-45 cm depth (Fig. 1 B).
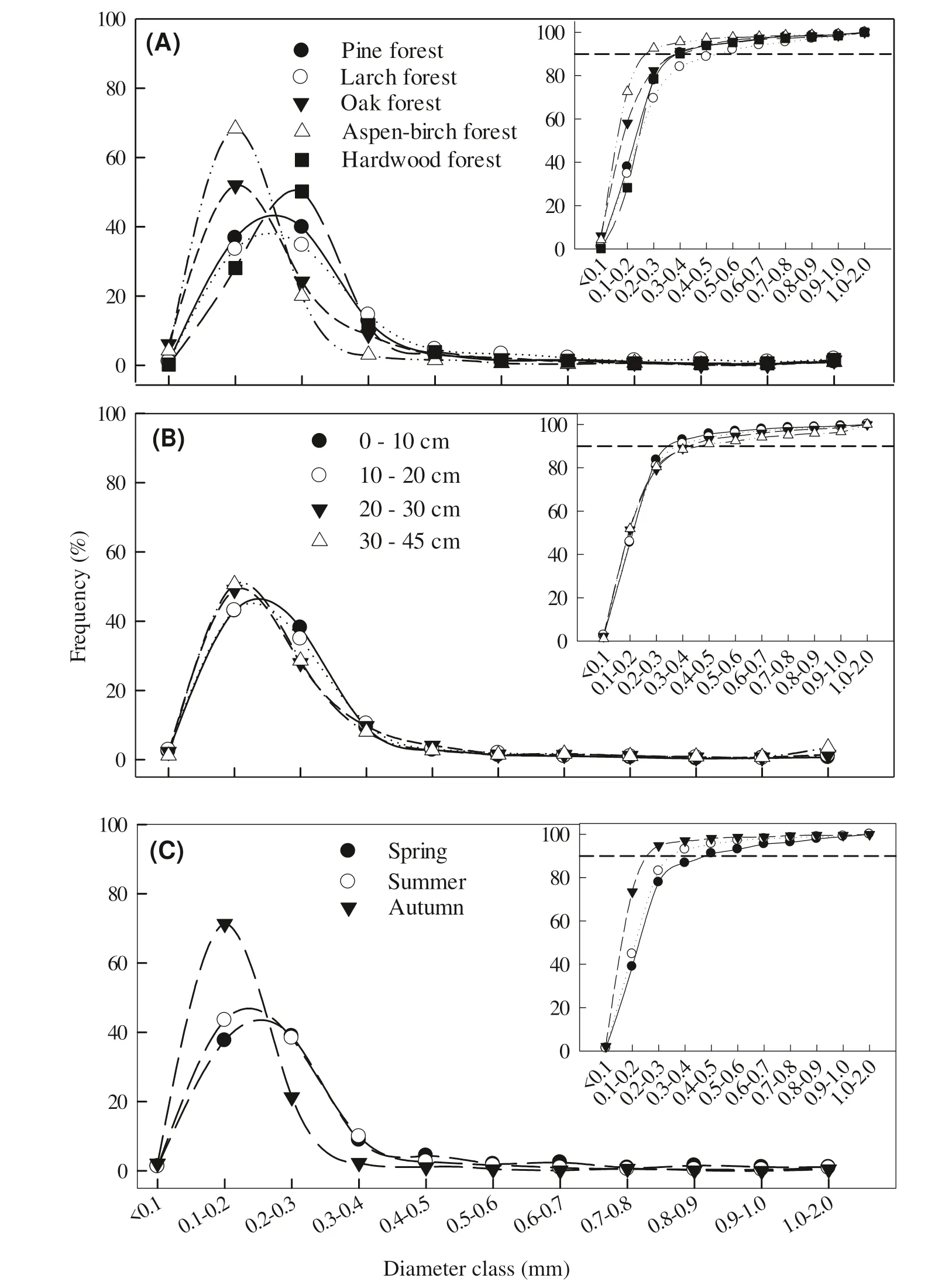
Fig. 1 Frequency of fine root numbers by diameter classes for the five forest types ( A), four soil depths ( B), and three growing season periods ( C). Inner panels show the sum of frequencies of fine root numbers with increasing diameters
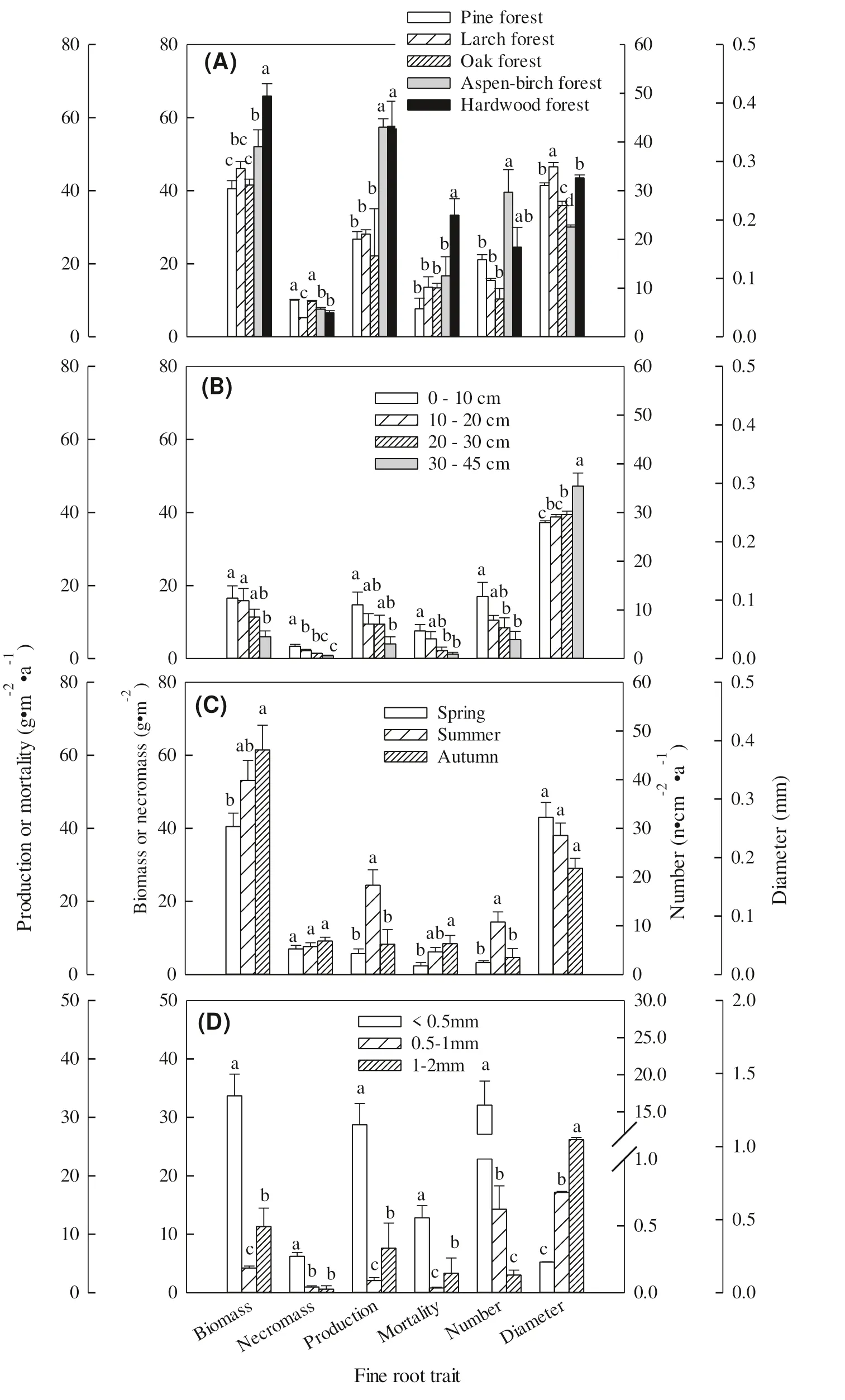
Fig. 2 Mean values of fine root biomass, necromass, production, mortality, number, and diameter among the forest types ( A), soil depths ( B), growing season periods ( C) and diameter classes ( D). The letters “a-d” denote, based on LSD tests at α = 0.05 level, signifi cant difference groups of means among the forest types ( n = 15), soil depths ( n = 20), seasons ( n = 15) and diameter classes ( n = 15). Error bars are standard deviation errors
Fine root diameters were significantly different among forest types and at soil depths but not among seasons (Fig. 2). The average fine root diameters for pine, larch, oak, aspen-birch and hardwood forests were 0.3, 0.3, 0.2, 0.2, and 0.3 mm, respectively. Conifers had thicker fine roots than broadleaved species (Fig. 2 A). Fine root diameter signifi -cantly increased with soil depth (Fig. 2 B), which the values varied from 0.2 to 0.3 mm. Although the means of fine root diameters did not differ significantly over the growing season, they decreased from the previous growing season (0.2-03, Fig. 2 C).
The number of fine roots was significantly different among forest types, at different soil depths, and in different seasons (Fig. 2). The means varied from 8 ± 2 cm -2 a -1 for the oak forest to 30 ± 5 cm -2 a -1 for the aspen-birch forest (Fig. 2 A). The numbers decreased from 13 ± 3 cm -2 a -1 in the 0-10 cm depth to 4 ± 2 cm -2 a -1 in the 30-45 cm depth (Fig. 2 B). The average number of fine roots produced in the spring, summer and autumn were 3, 11, and 4 cm -2 a-1, respectively (Fig. 2 C). The means in the < 0.5 mm, 0.5-1.0 mm and 1.0-2.0 mm diameter classes were 15.9, 0.6, and 0.1 cm -2 a -1 , respectively (Fig. 2 D).
The number of fine roots showed dissimilar distribution patterns along the soil profile (Fig. 3 A), decreasing as soil depth increased for the pine, oak, and hardwood forests. However, the numbers were in a sine-curve vertical pattern for the aspen-birch forest, while it was generally evendistributed for the larch forest across soil depths (Fig. 3 A). Peaks appeared at 0-10 cm and 20-30 cm for the aspenbirch forest, and 10-20 cm for the larch forest. The average frequencies of fine roots were 41.0%, 27.7%, 19.4% and 11.9% in the 0-10 cm, 10-20 cm, 20-30 cm, and 30-45 cm depth, respectively.
Fine root numbers had similar distribution patterns throughout the growing season, with a maximum in the summer (Fig. 3 B). The average frequencies of fine roots were 17.3%, 65.9%, 16.8% in the spring, summer, and autumn, respectively. There were similar distribution patterns in the three diameter classes, with maximum in the < 0.5 mm class (Fig. 3 C). The average frequencies of fine roots were 93.6%, 5.0%, 1.4% in the < 0.5 mm, 0.5-1.0 mm, and 1.0-2.0 mm diameter classes, respectively.

Fig. 3 Frequency of fine root numbers ( A- C), biomass ( D- F), necromass ( G- I), production ( J- L), and mortality ( M- O) in the five forest types by soil depth, growing season, and diameter classes
Fine root b iomass and necromass
The relative distribution of fine root biomass along the soil profile varied with forest type, while necromass decreased with increasing soil depth except for the larch plantation (Fig. 3 D, G). Both frequencies of biomass and necromass for the larch forest were relatively low in the upper 10 cm, peaked in the 10-20 cm depth, and declined afterwards. The frequencies of biomass for oak and hardwood forests decreased with increasing soil depth while for the pine and aspen-birch forests, they were relatively high at the soil surface, decreased at the 10-20 cm depth, peaked at the 20-30 cm depth, and then declined. Over the different soil depths, biomass frequency varied from 4.6 to 44.4%, while that of necromass ranged from 7.6 to 48.6%.
Both frequencies of biomass and necromass increased over the growing season for all forests (Fig. 3 E, H). The frequency of fine root biomass varied from 23.0 to 46.0%, while necromass ranged from 27.2 to 43.8%.
The frequencies of biomass for all forests peaked in the < 0.5 mm diameter class, and were relatively low in the 0.5-1 mm class, but necromass frequencies decreased with increasing diameter for all forests (Fig. 3 F, I). Biomass frequencies varied from 59.0-84.2%, 6.2-10.4%, and 6.5-31.9% for the < 0.5 mm, 0.5-1 mm, and 1.0-2.0 mm diameter classes, respectively, while necromass ranged from 74.7-82.7%, 7.0-5.5%, and 5.5-9.5%, respectively.
There were signifi cant Differences in biomass and necromass amounts among forest types, soil depths, diameter classes, and growing season ( P < 0.05) except for necromass amounts over the growing season ( P > 0.05, Fig. 2). Biomass amounts were 40.5, 46.0, 41.6, 52.1, and 65.9 g m-2for the pine, larch, oak, aspen-birch, and hardwood forests, respectively, while the corresponding necromass amounts were 10.0, 5.2, 9.7, 7.6, and 6.6 g m -2 (Fig. 2 A); biomass/necromass ratios were 4.0, 8.8, 4.3, 6.9, and 10.0, respectively. Average biomass and necromass amounts decreased with increasing soil depth and over the growing season, varying from 6.0-16.6 g m -2 and from 0.79-3.4 g m -2 over the soil profile, respectively. They ranged from 40.5-61.5 g m -2 and 7.0-9.2 g m -2 over the growing season, respectively (Fig. 2 B, C). The average biomass was 33.7, 4.2 and 11.3 g m -2 for the < 0.5 mm, 0.5-1 mm and 1.0-2.0 mm diameter classes, respectively, while the average necromass amounts were 6.2, 1.0 and 0.6 g m -2 , respectively (Fig. 2 D).
Fine root p roduction and mortality
The relative distribution patterns of fine root production and mortality along the soil profile were similar to those of the corresponding amounts of biomass and necromass, respectively (c.f., Fig. 3 D, G, J, M). The vertical patterns of production frequency varied substantially with forest type, varying from 3.8 to 57.5%. However, the frequency of mortality decreased with increasing soil depth, varying from 1.3 to 71.6%. Unlike biomass and necromass amounts, the frequencies of production and mortality over the growing season were different in terms of both pattern and divergence among the forest types (c.f., Fig. 3 E, H, K, N). Production of fine roots peaked in summer for all forests (66.3% average), whereas mortality maximized in autumn (mean of 48.4%). Both frequencies of production and mortality among the five forest types were highest in the < 0.5 mm class, ranging from 49.9 to 98.5% for production and 55.5-95.7% for mortality (Fig. 3 L, O).
Production and mortality of fine roots were significantly different among forest types, soil depths, diameter classes and seasons ( P < 0.05). Production was 26.7, 28.1, 22.1, 57.4, and 57.7 g m -2 a -1 for the pine, larch, oak, aspen-birch, and hardwood forests, respectively, while the corresponding fine root mortality was 7.7, 13.6, 13.4, 16.7, and 33.3 g m -2 a -1 (Fig. 2 A). The coniferous forests had less fine root production and mortality (27.4 g m -2 a -1 and 10.6 g m -2 a -1 ) than the broadleaved forests (45.7 g m -2 a -1 and 21.1 g m -2 a -1 ). Mean production and mortality decreased with soil depth, varying from 14.7 to 4.0 g m -2 a -1 and from 7.6 to 1.3 g m -2 a -1 , respectively. Overall, fine root production peaked in summer, with a maximum of 24.4 g m-2a -1 ; mortality, however, increased from 2.3 g m -2 a -1 in the spring to 8.4 g m -2 a -1 in autumn (Fig. 2 B, C). The mean production was 28.7, 2.1, and 7.6 g m -2 a -1 for the < 0.5 mm, 0.5-1 mm and 1.0-2.0 mm diameter classes, respectively, while average mortality was 12.8, 0.8 and 3.3 g m -2 , respectively.
Fine root t urnover
Fine root turnover (TR) were significantly different among the five forest types, soil layers and seasons for all three diameter classes ( P < 0.05). TR for the pine, larch, oak, aspen-birch, and hardwood forests in the < 0.5 mm class were 0.39 a -1 , 0.83 a -1 , 0.50 a -1 , 0.97 a -1 , and 0.75 a -1 , respectively, while for the 0.5-1.0 class they ranged from 0.21 a -1 for the oak forest to 0.49 a -1 for the hardwood forest, and for the 1-2 mm class, from 0.07 a -1 for the oak forest to 0.26 a -1 for hardwood forest (Fig. 4). Turnover rates in the 0-10 cm depth were 1.0 a -1 , 0.79 a -1 and 0.49 a -1 for the < 0.5 mm diameter class, 0.5-1.0 mm class and 1.0-2.0 mm class, respectively, and approximately 75.4%, 115.2% and 65.9% greater than the average for the rest of the soil (0.57 a -1 0.36 a -1 and 0.30 a -1 ). TR peaked at 1.09 a -1 , 0.61 a -1 and 0.20 a -1 for the < 0.5 mm class, 0.5-1.0 mm class and 1.0-2.0 mm class in summer, more than twice the mean of other seasons. The average turnover rates of forest types, soil depths and seasons in the < 0.5 mm diameter class was 0.69 a -1 , 0.68 a-1and 0.65 a -1 , respectively, approximately 109.1%, 172% and 92.6% greater than in the corresponding 0.5-2.0 mm class.

Fig. 4 Fine root turnover rates by forest type, soil depths, and growing season. The letters “a, b” denote, based on Duncan tests at α = 0.05 level, signifi cant difference groups of means among the forest types, soil depths, and growing season. Error bars are standard errors ( n = 15, n = 20, and n = 15, respectively)

0.70** 0.27 0.65** 0.57* 0.35 0.93** 0.51* 0.71** 0.39 K A (mg kg -1 ) - 0.09 - 0.21 P A (mg kg -1 ) 0.33 0.32 0.17 0.002 0.30 0.34 0.57* 0.35 0.37 0.73** - 0.30 Table 4 Correlation coeffi cients between fine root traits and soil properties obtained by pooling the data across forest types, soil depths and seasons 0.44* 0.42 0.57* 0.14 0.21 0.20 K (mg g -1 ) 0.44** 0.22 0.35 - 0.22 - 0.02 0.49* 0.61** 0.44* 0.18 0.56* 0.38 P (mg g -1 ) 0.68** 0.46* 0.65** - 0.43 - 0.14 0.01 0.37 0.50* 0.41 0.16 0.75** 0.26 NO 3 - (mg kg -1 ) - 0.08 0.64** 0.20 - 0.33 NH4 + (mg kg -1 ) 0.60** 0.03 0.73** 0.46* 0.58* 0.35 0.10 - 0.34 - 0.20 - 0.06 - 0.08 0.67** 0.28 0.67** - 0.04 0.38 0.51* 0.54* 0.88** 0.46* 0.75** W 5 (g H 2 O g -1 soil) N (mg g -1 ) - 0.34 0.62** 0.28 0.70** 0.04 0.45* 0.34 0.87** 0.42 0.72** 0.45* - 0.3 T 5 (°C) 0.38* 0.43* 0.42* 0.40* - 0.08 0.56** 0.65** 0.38* 0.50** 0.43* - 0.16 Diameter class (mm) < 0.5 0.5-2 < 0.5 0.5-2 < 0.5 0.5-2 < 0.5 0.5-2 < 0.5 0.5-2 < 2 Variable Biomass (g m -2 ) Necromass (g m -2 ) Production (g m -2 a -1 ) Mortality (g m -2 a -1 ) Number (n cm -2 a -1 ) Diameter (mm) T 5 , W 5 , N, NH 4 + , NO 3 - , P, P A , K and K A at 5-cm depth are soil temperature soil water content, nitrogen, ammonium and nitrate concentrations, phosphorus and available phosphorus concentrations, and potassium and available potassium concentrations, respectively *,**Signifi cance levels at α = 0.05 and α = 0.01, respectively. n = 23 for T 5 , n = 15 for W 5 , N, NH 4 + , NO 3 - , P, P A , and K A
Fine root traits related to soil properties
Fine root diameter was negatively correlated with most of the soil property variables but the correlation was insignifi -cant. For the < 0.5 mm diameter class, fine root numbers, biomass, necromass, production, and mortality were all positively correlated with T5(soil temperature), W5(soil water), KA(available potassium), P (phosphorous), and N (nitrogen), with correlation coeffi cients varying from 0.38 to 0.93 (Table 4). For the 0.5-2.0 mm diameter class, only T5significantly infl uenced the fine root number, biomass, mortality and production; while the fine root number was also infl uenced by W5, N, P and PA. Necromass levels were not significantly correlated with all the variables (Table 4).
Discussion
Fine root dynamics and turnover rates by diameters
Fine root diameter is an important morphological index refl ecting tree characteristics for resource acquisition (Eissenstat and Yanai 2002). Our data show that more than 90% of the fine roots throughout the soil profiles in the five forest types and over the growing season were < 0.5 mm in diameter (Fig. 1). This is in agreement with previous studies (Hendrick and Pregitzer 1992; Wells and Eissenstat 2001; McCormack et al. 2015). In addition, the high frequency of this diameter class accounted for more of the biomass, necromass, production and mortality than the low frequency of the 0.5-2.0 mm class. More frequent < 0.5 mm fine roots are better correlated with soil properties than the 0.5-2.0 mm diameter roots (Table 4). These results suggest that it would be inaccurate to treat all roots < 2.0 mm as a functionally homogenous group, and also support the argument that the conventional category of fine roots (i.e., < 2 mm diameter) can be divided into morphological and physiological subgroups for various tree species (Wells and Eissenstat 2001; Pregitzer et al. 2002; Guo et al. 2004; Wang et al. 2006b; McCormack et al. 2015). McCormack et al. ( 2015) suggested that the broadly defined fine-root group should be split into absorptive fine roots and transport fine roots. Absorptive fine roots are closely related to resource acquisition and uptake, whereas transport fine roots have primarily transport functions. This approach could enable comparisons among functionally similar roots; however, in this study, the identifi cation of root traits to determine functional divisions was inconvenient in the mixed forest. In this study, the fine root frequency method was similar with the fine root functional classifi cation in number and diameter (McCormack et al. 2015), and still allowed meaningful comparisons of biomass, necromass, production and mortality across forest types. Moreover, it can reduce sample processing times and avoid the need to identify individual root functions.
Accurately estimating root turnover rates is essential for the evaluation of the role of fine roots in the cycling of water, nutrients, and carbon in terrestrial ecosystems. Our data show that the turnover rates decreased with increasing fine root diameter classes (Fig. 4). The data also show that turnover rates were different between the more frequent < 0.5 mm diameter fine roots and the 0.5-2.0 mm diameter roots. The mean turnover rates by forest type, soil depth and seasons for < 0.5 mm fine roots were 2.09, 2.72 and 1.93 times higher than the turnover rates for the 0.5-2.0 mm roots (Fig. 4). This suggests that it was incorrect to assume all fine roots < 2 mm had the same rates of turnover. In reality, only the more frequent < 0.5 mm diameter fine roots turn over more quickly and comprise most of the biomass and production of roots < 2.0 mm in diameter (Fig. 2). The mean turnover rates by forest types for roots < 2.0 mm in diameter was 0.54 a -1 , 28.5% lower than for < 0.5 mm diameter roots (0.68 a -1 ).
Fine root dynamics and turnover rates by forest types
Fine root diameter, number, biomass, necromass, production, and mortality were all significantly different among the five forest types in the same geographical region (Figs. 1 A, 2 A). This is similar to previous studies (Hendrick and Pregitzer 1993; Steele et al. 1997; Wang et al. 2006b; Quan et al. 2010). These Differences in fine root properties may be attributed to the genetics of the species and their adaptation to environmental changes (Comas and Eissenstat 2004; Eissenstat et al. 2015; Zadworny et al. 2016). Compared with the coniferous plantations, the broadleaved naturally regenerated forests had a greater frequency of thin fine roots (Fig. 1 A) and root numbers, resulting in greater biomass, necromass, production and mortality (Fig. 2 A), supporting our first hypothesis. This is also consistent with the general morphological distinction between angiosperms and gymnosperms (López et al. 2001; Pregitzer et al. 2002; Wang et al. 2006b). Our data also shows that fine root number, biomass, necromass, production, and mortality were all positively correlated with soil temperature, water content, available potassium, phosphorous and nitrogen (Table 4). The aspenbirch and hardwood forests normally occur at mid- and toeslopes with moist, fertile soils (Wang 2006), probably leading to greater fine root numbers, biomass and production. In contrast, the oak forest is found on upper slopes with arid infertile soils, resulting in lower root numbers, biomass and production (Fig. 2 A).
There was no signifi cant relationship between fine root diameters and soil properties (Table 4). This may be associated with mycorrhizal colonization. Fine roots of coniferous species are colonized by ectomycorrhiza fungi, while those of broadleaved species are colonized by endomycorrhiza fungi (Brundrett 2002; Pregitzer et al. 2002). Additionally, co-occurring species in a same forest may have an asynchronous phenology and nutrient adaptive strategy by fine roots (Burke and Raynal 1994; Burton et al. 2000). These factors may contribute to the greater diameters of the former (Comas and Eissenstat 2004; Wang et al. 2006b) and mask the impact of soil properties on fine root diameters (Table 4). The negative relationship between fine root diameter and soil properties examined in this study indicates that fine roots can change their morphology and surface area to acquire soil resources more effi ciently.
The fine root turnover rates in this study (0.39-0.98 a-1for < 0.5 mm diameter) were within the range for temperate forests (0.1-2.0 a -1 ) (Gill and Jackson 2000). There were signifi cant Differences in turnover rates among the five forest types for both < 0.5 mm and 0.5-2.0 mm root diameter classes, possibly because of signifi cant inter-stand variations (Fig. 4), suggesting that forest type is an important consideration for accurately determining turnover rates of fine roots.
Several possible factors may contribute to inter-stand variations in turnover rates (Gill and Jackson 2000). First, the genetic characteristics of tree species may partly explain variability (Eissenstat and Yanai 2002). Fast-growing species (e.g., aspen, birch, larch) usually have higher turnover rates than slower growing species (e.g., oak, pine) (Eissenstat and Yanai 2002). The coniferous forests had lower average turnover rates (0.61 a-1for fine roots < 0.5 mm diameter) than broadleaved forests (0.74 a -1 for fine roots). This is in agreement with other studies (Vogt and Bloomfi eld 1991; Coleman et al. 2000; Matamala et al. 2003). Secondly, soil resource availability infl uenced fine root production and mortality (Table 4), and thus aff ected turnover rates (Hendrick and Pregitzer 1993, 1997; Lee and Jose 2003), which altered the species-associated trend described above. For example, some studies indicate that coniferous forests have higher turnover rates than broadleaved forests on various boreal sites (Ruess et al. 1996; Steele et al. 1997). Among the broadleaved forests in this study, the hardwood and aspen-birch forests had greater turnover rates than the oak forest, possibly due to their Differences in soil resource availability (Fig. 4). Thirdly, multi-species mixed forests may possess greater variability in stand-based turnover rates than monoculture plantations because of species divergence in growth characteristics and responses to environmental conditions. Fourthly, the fine root classifi cation method may result in diverse turnover rates. In this study, for fine root diameters < 0.5 mm, the aspen-birch forest had the highest turnover rates and the pine plantations had the smallest. However, the hardwood forest had the highest turnover rates and the oak forest had the smallest for fine root diameters 0.5-2.0 mm. Fifthly, measuring uncertainty may also introduce bias in turnover rate estimates. In this study, as suggested by Johnson et al. ( 2001), MR images were collected every 2 weeks. However, biomass and production of fine roots were considerably greater than the necromass and mortality for the five forest types (Fig. 2 A), implying belowground carbon accumulation or/and fine root decomposition between measuring periods. In spite of this, the turnover rate was less than 1 a -1 (i.e., the fine root longevity was greater than 1 a), and biomass was greater than necromass, especially for the aspen-birch and hardwood forests (Fig. 2 A), again indicating that some carbon loss from fine root decomposition occurred. These results suggest that turnover rates in this study may be underestimated for some forests.
Fine root dynamics and turnover rates along the soil profile
Most thin (< 0.5 mm) fine roots appeared in the surface soil layers for all forests, while those with larger diameters (0.5-2.0 mm) occurred at deeper layers (Fig. 2 B). This finding, similar to other studies (Wells and Eissenstat 2001; Eissenstat and Yanai 2002) may refl ect plant cost-benefit principles. Shallower occurring thin fine roots with greater surface area may be more capable of absorbing nutrients at surface soil layers where more nutrients are available; in contrast, deeper, thicker fine roots can penetrate into lower soil layers where nutrients are relatively defi cient, soil bulk density is higher, and water supply may be more stable (Eissenstat 1992; Eissenstat et al. 2000; Hutchings and John 2003; Brassard et al. 2009).
Both relative and absolute distribution of the numbers of fine roots, biomass, necromass, production of fine roots, and fine root mortality decreased with soil depth (Figs. 2, 3), in agreement with previous research (Powell and Day 1991; Hendrick and Pregitzer 1996). Few studies, however, quantify fine root dynamics or characteristics at the forest level, in spite of the fact that these patterns of distribution seem to be determined (Fahey and Hughes 1994; Hendrick and Pregitzer 1996, 1997; Ruess et al. 2003; Baddeley and Watson 2004). Hendrick and Pregitzer ( 1996) reported that 44% of the production and 40% of the mortality of fine roots were concentrated in the upper 20-cm depth in a northern hardwood forest. Our data shows that the upper 20-cm soil layer contained relatively high percentages of fine root production (55.4-72.6%) and mortality (64.8-95.9%) (Fig. 3 J, M). Although there was a general decreasing trend in fine root number, biomass, necromass, production, and mortality with soil depth, the values and depth of maximum occurrence differed among the forest types (Fig. 3). This highlights the signifi cance of understanding various belowground carbon allocation patterns for different forest types in modeling forest carbon and nutrient cycles (Jackson et al. 1997).
In addition toforest types, the vertical distribution of fine roots may also be associated with the soil environment and resource availability (Canadell et al. 1996; Hendrick and Pregitzer 1996; Jackson et al. 1996; Rytter and Hansson 1996; Burton et al. 2000; Quan et al. 2010). The fact that most fine root traits were closely related to soil temperatures, soil water contents and nitrogen (Table 4), which refl ected the impact of site conditions on fine root dynamics. Several studies show that the deeper the soil, the lower the turnover rate or the longer lifespan the fine roots have (Kosola et al. 1995; Burton et al. 2000; Johnson et al. 2001; Wells et al. 2002; Anderson et al. 2003). Pooling data across the five forest types, showed that turnover rates decreased with soil depth for all diameter classes (Fig. 4). The fine roots in the surface layers have thinner diameters and large surface area (Figs. 2 B, 3), and maintain active metabolism facilitated by warm temperatures and high nitrogen availability (Table 4). The increased maintenance respiration consequently results in high turnover rates to improve carbon use efficiency (Burton et al. 2000; Gill and Jackson 2000; Majdi and Öhrvik 2004). In contrast, fine roots at deeper layers have thicker diameters and low possibility of pathogen invasion, leading to prolonged longevity or lower turnover rates (Gill and Jackson 2000; Wells and Eissenstat 2001; Baddeley and Watson 2005).
Fine root c haracteristics and turnover rates over the growing season
In spite of insignifi cant Differences in fine root diameters over the growing season, there was a trend in diameter reduction as the growing season began (Fig. 2 C). This may be attributed to the course of development of fine roots (i.e., a low order root in spring might become a higher order root in September), to cortical senescence or cell death (Liljeroth 1995; Wang et al. 2006b), or to changes in tissue density over the growing season (Eissenstat and Yanai 1997; Pregitzer et al. 2002). However, some studies have reported that autumn-developed fine roots were thicker so as to store carbohydrates for the coming year’s growth and to prepare for winter (Eissenstat and Yanai 1997; Fitter et al. 1998; Pregitzer et al. 2002; Pregitzer 2003). Our data also shows that, unlike other forests, the oak forest had the largest diameter of fine roots in autumn (data not shown). Further monitoring of fine root diameters by species and seasons are needed to discern possible homeostatic mechanisms for diameter seasonality.
Previous studies in temperate forests have shown different seasonal patterns of fine root production, such as unimodal with a maximum in spring (Hendrick and Pregitzer 1993, 1997; Lee and Jose 2003) or in summer (Burke and Raynal 1994; Rytter and Hansson 1996; Ruess et al. 1998, 2003; Brassard et al. 2009) or with a bimodal peak occurring in spring and autumn (Hendrick and Pregitzer 1996; Dress and Boerner 2001; Son and Hwang 2003). In this study, more than 66% of fine root number and production occurred in summer (Figs. 2 C, 3 B, K). The likely reasons for this summer peak are that: (1) favorable temperatures, abundant precipitation and available soil resources accelerated the growth of fine roots (Table 4) (Pregitzer et al. 2000; Chapin et al. 2002; Quan et al. 2010); and, (2) maximum leaf area in summer provided an abundant supply of carbohydrates for fine root growth (Vogt et al. 1996; Burton et al. 1997; Pregitzer et al. 2000; Quan et al. 2010). Nevertheless, possible mechanisms for other seasonal patterns of fine root numbers and production are lacking.
Both relative and absolute distributions of fine root biomass, mortality, and necromass showed progressively increasing trends over the growing season (Figs. 2, 3). This might be associated with aboveground phenology (Fahey and Hughes 1994; Pregitzer et al. 2000) and/or Soilconditions, especially in temperate forests (Pregitzer et al. 1993; King et al. 1999). There is normally a time lag between fine root production and biomass accumulation (Dress and Boerner 2001). It is thus possible to attain peak biomass in autumn, which may be benefi cial for storing non-structural carbon for the growth in the coming spring. In the late growing season, when temperatures have decreased and leaves begin tofall, a large quantity of fine roots died (Farrar and Jones 2000), possibly following the carbon cost-benefit model (Eissenstat and Yanai 1997), accordingly mortality increased (Fogel 1983; Ericsson et al. 1996; Ruess et al. 1998). The increasing mortality and low decomposition rates at low temperatures resulted in maximum fine root necromass in autumn (Silver and Miya 2001).
Our data shows that the rate of turnover of fine roots peaked in summer for all diameter classes (Fig. 4). Because environmental conditions in summer are benefi cial for biomass accumulation, production and mortality (Table 4) (Burton et al. 1997; Eissenstat et al. 2000; Pregitzer et al. 2000), fine root lifespan shortened, resulting in high turnover rates (Ryan et al. 1996; Burton et al. 2000; Pregitzer et al. 2000).
Conclusion
Patterns of fine root numbers, biomass, necromass, production, mortality, and turnover rates varied in the Chinese temperate forest types, with soil depths, over the growing season and with diameter classes. These forest-level patterns may be associated with the genetic characteristics of the tree species and soil properties. The findings highlight the importance of forest composition and root diameters in quantifying fine root dynamics and turnover rates, and also emphasize the necessity of considering the infl uence of both forest types and environmental conditions on fine roots when measuring and modeling carbon and nutrition cycles. A better understanding of fine root dynamics and belowground carbon allocation needs to couple belowground with aboveground processes and combine morphological monitoring with physiological measurements in future studies.
AcknowledgementsThe Maoershan Forest Ecosystem Research Station provided fi eld logistic support for the research.
杂志排行

Journal of Forestry Research的其它文章
- Do increasing respiratory costs explain the decline with age of forest growth rate?
- At what carbon price forest cutting should stop
- Mapping the risk of winter storm damage using GIS-based fuzzy logic
- Effects of seed moisture content, stratification and sowing date on the germination of Corylus avellana seeds
- Comparison of seed morphology of two ginkgo cultivars
- De novo assembly of the seed transcriptome and search for potential EST-SSR markers for an endangered, economically important tree species: Elaeagnus mollis Diels