Effects of seed moisture content, stratification and sowing date on the germination of Corylus avellana seeds
2020-05-19EliasPipinisAthanasiosStampoulidisEliasMiliosKyriakiKitikidouSofiaAkritidouStavroulaTheodoridouKalliopiRadoglou
Elias Pipinis · Athanasios Stampoulidis · Elias Milios · Kyriaki Kitikidou ·Sofia Akritidou · Stavroula Theodoridou · Kalliopi Radoglou
Abstract For Corylus avellana, pericarp permeability,seed desiccation tolerance,and the effect of warm and cold moist stratification combinations on germination and of outdoor sowing date on dormancy breaking and seedling emergence were investigated. Seeds with (intact fruit nut)and without the pericarp were subjected to an imbibition test, and water uptake was measured. After nuts had been stored for 5 months at 3-5 °C, seeds and desiccated seeds were cold-stratified at 3-5 °C (for 0, 1, 2 or 3 months),then tested for germination. Nuts were warm-stratified at 20-25 °C(for 0,1 or 2 months),then cold-stratified(for 0,1,2 or 3 months)and tested for seed germination.The nuts were sown outdoors on three dates in the same year of their collection: 15 September (immediately after collection), 2 November,and 17 December,and emerged seedlings were counted the following spring. The pericarp was not a barrier to water uptake by the seed; however, the seeds imbibed water faster and had more mass when the pericarp was removed. Seeds tolerated desiccation; germination percentage was not reduced when seed moisture content was reduced to 8.71%. Germination of seeds without the pericarp (regardless of the moisture content) was the highest when the seeds were cold-stratified for 2 months.When seeds with the pericarp were tested for germination,2 months of warm stratification before a 2-month cold stratification maximized seed germination. For outdoor sowing of nuts,sowing date affected percentage of seedling emergence the following spring; early autumn sowing(immediately after collection) resulted in the best emergence.
Keywords Corylus avellana · Desiccation tolerance ·Dormancy · Pericarp · Water uptake
Introduction
The genus Corylus (hornbeam) includes about 15 species of deciduous, large shrubs (rarely small trees) in the temperate regions of North America, Europe and Asia (Barbour and Brinkman 2008).The species Corylus avellana L.(European hazel) is native to Europe and west Asia(Browicz 1982). In Greece, C. avellana is a common species and usually found in mountainous and submountainous regions of the continental part of the country(Boratynski et al. 1992). It is an economically important species, cultivated for its edible fruits (nuts). Furthermore,the ecological significance of C. avellana in a forest ecosystem is great,due to the forage(nuts)for wildlife.As a result, the species has to be propagated in nurseries.
In reforestation or genetic breeding programs, the massive propagation of seedlings with broad genetic diversity that is needed can be only achieved with sexual propagation(Macdonald 2006).Although this procedure is simple,seed dormancy is one of the biggest hurdles to overcome and has often been a challenge. Usually, the germination unit of C. avellana is a seed enclosed by a woody hard pericarp, known as nut. According to Bradbeer et al.(1978), the pericarp, testa, and embryo are involved in the delayed germination of C. avellana seeds. The pericarp could be water impermeable, permeable to water but mechanically resistant to expansion of the embryo, or contain substances that inhibit germination (Baskin and Baskin 1998). According to Jarvis (1975), removal of the pericarp and testa stimulate germination of freshly harvested seeds. Dormancy of freshly harvested seeds is imposed by inhibitors in testa and pericarp and may also be the result of mechanical resistance of the pericarp (Bradbeer 1968).However,there is no published information on the water permeability of the pericarp in C. avellana nuts.Furthermore, Jarvis (1975) and Bradbeer (1968) demonstrated that during the dry storage of C. avellana seeds,inhibitors in the pericarp and testa are carried into embryonic axis and induce a deeper state of dormancy. In this case,cold stratification and gibberellic acid application are used to overcome dormancy in seeds after the pericarp is removed(Jarvis and Wilson 1977;Bradbeer et al.1978).
Although a number of studies have been carried out on the nature of dormancy and its release in C.avellana seeds without the pericarp, no attention has been given to overcoming dormancy of the true seed enclosed by the hard pericarp.Usually,for practical reasons,the true seed is not extracted from the hard pericarp and germinates from within the pericarp. In this case, an appropriate treatment has to be applied to overcome the dormancy imposed by the pericarp. Laboratory studies have shown that in a number of species with seeds enclosed by a hard pericarp,dormancy break and germination are promoted by a warm stratification before a cold stratification (Tylkowski 1992;Morgenson 2000; Chen et al. 2007). The beneficial effect of warm stratification on seed germination has been attributed to a decrease in pericarp mechanical resistance(Persson et al.2006).Alternatively,this treatment of seeds(with pericarp) can be achieved by outdoor sowing in the nursery at a time when the natural environment provides the necessary conditions to overcome dormancy(Hartmann et al. 2011). According to Baskin and Baskin (1998),temperature is the major environmental factor responsible for changes in the dormancy states of seeds.Seeds of many species in the temperate zone require exposure to low temperatures for dormancy break; consequently, they become non-dormant during the winter and germinate the following spring. In other species, seeds with a fully developed embryo require exposure to summer, followed by autumn and winter temperatures before they germinate the next spring (Baskin and Baskin 1995; Baskin et al.2002). When the required sequence of temperature conditions for seed dormancy release is not known and the seeds are exposed to winter temperatures only, dormancy break will not occur, and poor germination will result in increased production costs for the nursery.
Nurseries must also store seeds properly for maximum viability to ensure supply when they are needed for massive seedling propagation in a reforestation programme.Moisture content is the most important factor affecting seed longevity during the storage. Based on their response to desiccation, seeds in general have been classified as orthodox (desiccation tolerant), recalcitrant (desiccation sensitive) (Roberts 1973), or suborthodox (intermediate)(Ellis et al. 1990). However, seeds of C. avellana have been classified variously as recalcitrant(Heit 1967;Roberts 1973), suborthodox (Bonner 1986) and orthodox (Normah et al. 1994; Gosling 2007; Michalak et al. 2013). Furthermore, Hartmann et al. (2011) classified the seeds of Corylus species as recalcitrant and recommended moist storage at low temperature (0-10 °C).
The general aim of the present study was to investigate dormancy breaking and germination requirements of C.avellana seeds enclosed by the pericarp, under laboratory and nursery conditions, to develop a reliable propagation protocol. The specific objectives of this research were to(1)examine the effect of pericarp on imbibition of the seed,(2)evaluate the impact of desiccation on seed germination,(3) describe the effects of warm and cold moist stratification treatment combinations on germination of seeds enclosed by the pericarp under laboratory conditions, and(4)to compare the effects of different outdoor sowing dates on dormancy breaking and seedling emergence of seeds enclosed by the pericarp.
Materials and methods
Seed collection
Fruits of C. avellana (nuts; the seed is enclosed by a hard pericarp)were collected from the ground on 10 September 2015 after their dispersion from many individuals growing in natural conditions in northeastern Greece (41°12′58′′M,24°38′55′′E, 123 m a.s.l.). After collection and removing visibly damaged nuts,the nuts were dried on filter papers in the laboratory for 2 days, and then stored in sealed glass containers at 3-5 °C until used in experiments the next February.
Water uptake
To determine the effect of the pericarp on permeability to water, an imbibition test was conducted at room temperature on four replicates of 30 nuts and 30 extracted seeds (the pericarp was removed after the nut was cracked in a vise). The total sample mass of each replicate (in total 8 replicates) was weighed, then the samples were immersed in distilled water in a glass(three volumes of water for each volume of seeds or nuts).After soaking for 3,6,12,24,36,and 48 h, then at 24-h intervals for the following 8 days,the seeds or nuts of each replicate were removed from water, quickly surface-dried with paper and then weighed.The percentage water uptake (Wp) was determined according to the following equation:
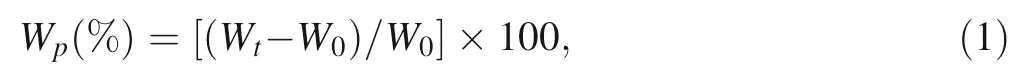
where Wtis the total seed or nut mass at time t, W0is the initial total seed or nut mass at the start of the experiment(time 0) (Li et al. 1999).
Effect of moisture content on seed germination
After nuts had been stored for 5 months, the moisture content of true seeds (initial moisture content; IMC) of collected nuts was determined according to the rules of the International Seed Testing Association (ISTA 1999).Afterwards, 420 randomly selected seeds (with out the pericarp)were subjected to desiccation in an oven at 35 °C.Twenty seeds that were randomly selected from the aforementioned 420 seeds were weighed individually before and during the desiccation at 1-h intervals until their moisture content(MC)was reduced to <10%.For each of the 20 seeds, the MC was calculated as:

where IMCis the initial moisture content of seeds,W0is the seed mass before the desiccation, Wtis the seed mass at interval t.
When the desired moisture content in the 20 seeds was achieved (after 7 h of drying) the desiccation procedure was stopped and the reduced moisture content of the 420 seeds was checked according to the rules of ISTA (1999).The desiccated seeds and 400 seeds with the initial moisture content were mixed with moist, sterilized river sand,placed in different plastic containers and cold-stratified for 0, 1, 2 and 3 months in a refrigerator at 3-5 °C.
Effect of stratification on germination of seeds enclosed by the pericarp
After 5 months of storage, 400 nuts of C. avellana were mixed with moist sterilized river sand in each of three plastic containers and warm-stratified for 0,1,or 2 months at 20-25 °C. At the end of each warm stratification (WS)period, a plastic container was placed in the refrigerator(3-5 °C)for cold stratification(CS)of the seeds for 0,1,2 or 3 months. In total, 12 pretreatments (WS and CS combinations) were applied.
Germination test
In each of the aforementioned experiments, at the end of the CS, a random sample of 100 seeds or nuts was taken from each plastic container,dusted with fungicide(Captan)and randomly placed in four plastic Petri dishes (25 seeds per Petri dish). For each pretreatment, there were four replications of 25 seeds.The seeds and nuts were placed on sterilized river sand, moistened with distilled water, in 12-cm plastic Petri dishes.The Petri dishes were randomly arranged on the shelves of a growth chamber at 20 °C for 16-h dark and 25 °C for 8-h light. Distilled water was added as needed. Germinated seeds were counted each week for 9 weeks. A seed with at least 2-mm-long radicle was considered to be germinated (ISTA 1999). For each treatment, the germination percentage (GP) and the mean germination time(MGT)were calculated as the average of the four replications. The MGT was calculated for each replication per treatment as

where n is the number of seeds that germinated on day D,and D is the number of days counted from the beginning of the test (Ellis and Roberts 1981).
Effect of sowing date on seedling emergence
Nuts were sown on three dates in the same year of their collection:15 September(immediately after collection and drying of nuts),2 November,and 17 December 2015.Nuts were sown outdoors in an area close to the collection site.On each sowing date,24 nuts were sown by hand in each of five plastic containers (Quick pot T/16, 24 cavities, 1 for each nut, cavity volume 330 cm3, filled with a 4:1 (v/v)mixture of peat and perlite). Nuts were carefully covered with sand about twice the size of the nut. The containers were then watered and covered with a plastic net until the next spring to protect nuts from predators(bird,rodent)and heavy rainfall and to maintain moisture in the substrate.Attention was given to watering to avoid drying or excess moisture in the substratum. In autumn, the watering frequency and duration depended on the weather conditions,whereas the containers were not watered in the winter.The next spring, more than one watering per day was required to maintain the proper moisture for seed germination.After the first seedling emerged in early March, emerged seedlings (i.e., the epicotyl was above the soil surface) in each container were counted at weekly intervals until the end of May.
Temperature conditions during the experiment(September 2015 to May 2016) are shown in Fig. 1. Air temperature data were drawn from the closest meteorological station of the National Observatory of Athens in Xanthi.
Statistical analyses
For all experiments, a completely randomized experimental design was used. The germination percentages as well as the percentages of seedlings which emerged from nuts sown were arcsine square-root transformed before analysis(Snedecor and Cochran 1980). The transformed data and the MGT data were checked for normality and homogeneity of variances and then analyzed by a one-way ANOVA. Comparisons of the means were made using the Duncan test (Klockars and Sax 1986). All statistical analyses were carried out using SPSS v. 21.0 software (SPSS,Inc., USA).
Results
Water uptake
Seeds of C. avellana without the pericarp imbibed water rapidly, increasing by 23.96% of their mass after 3 h(Fig. 2).Nuts imbibed less water;after 3 h,their mass only increased by 4.66%. After 24 h in water, the imbibition percentage of seeds was 51.52% and 11.73% for nuts. In seeds,the maximum rate of water imbibition was recorded during the first 24 h of immersion. After 10 days in water,the imbibition percentage of seeds had increased to 58.16%and to 27.77% for nuts.
Effect of moisture content on seed germination

Fig. 1 Mean, mean minimum and mean maximum air temperature values for each 15-day period from September 2015 to May 2016
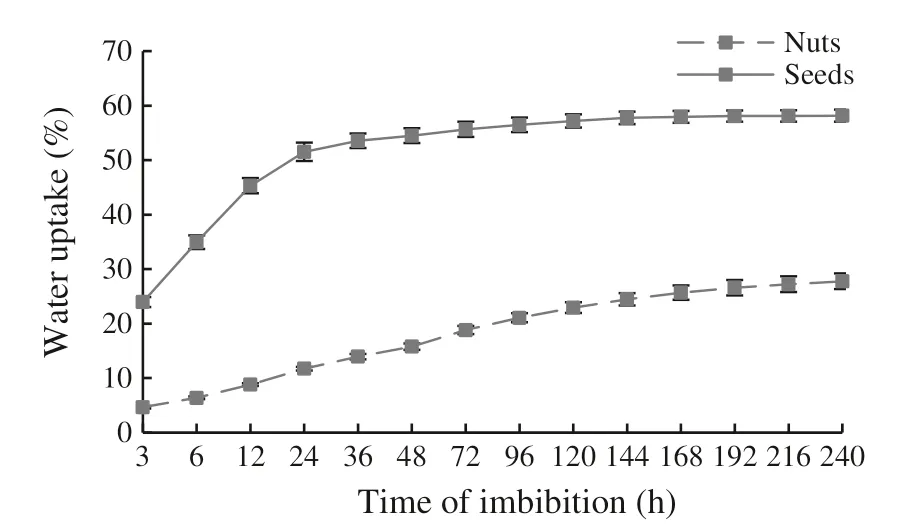
Fig. 2 Mean percentage (± SD) increase in mass over time of C.avellana seeds and nuts immersed in distilled water
In both level of MC of seeds, a CS period longer than 2 months was not used because at the end of the second month of CS, seeds had begun to germinate. After 5 months of storage in sealed glass containers at 3-5 °C,the MC of seeds was 20.73%(± 0.63),whereas the MC of seeds which were desiccated in an oven at 35 °C for 7 h was 8.71% (± 0.57) (Table 1).
For each CS period(0,1 and 2 months), the GP did not significantly differ between the 20.73% MC seeds and 8.71%MC seeds,nor did MGT(Table 1).The reduction in MC did not affect seed germination. Furthermore, regardless of the MC level, increasing the CS period to 1 and 2 months significantly increased GP and reduced MGT.
Effect of stratification on germination of seeds enclosed by the pericarp
None of the C. avellana nuts subjected to only 1 month of CS germinated, whereas an increase in the CS to 2 or 3 months resulted in low GPs (34 or 36%, respectively).Warm stratification significantly improved the GP. In nuts that had been warm-stratified for 1 month, a 3-month CS significantly improved the GP(65%)and reduced the MGT(10.96 days). However, increasing the WS to 2 months with a 2-month CS resulted in entirely breaking of dormancy (Table 2).
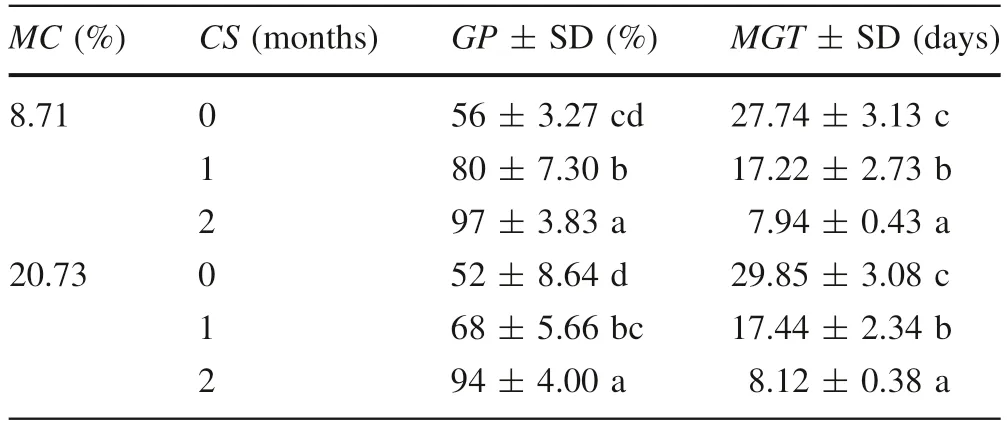
Table 1 Effects of seed moisture content(MC)and cold stratification(CS) on germination percentage (GP) and mean germination time(MGT) of Corylus avellana seeds
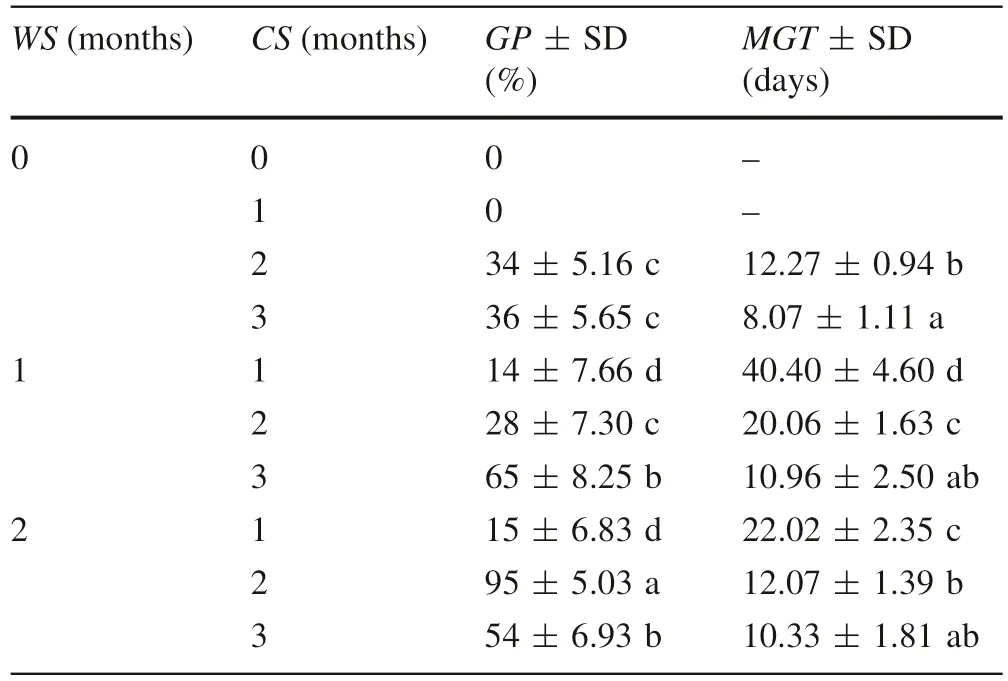
Table 2 Effect of warm stratification (WS) following by cold stratification (CS) on germination percentage (GP) and mean germination time (MGT) of Corylus avellana seeds enclosed by the pericarp
Effect of sowing date on seedling emergence
Sowing date significantly affected seedling emergence in the spring after sowing (Fig. 3). The highest percentage of seedling emergence was obtained when nuts were sown on 15 September (83.33%). Seedling emergence was lowest for nuts sown on 17 December (44.17%).
Discussion
True seeds and the nuts of C. avellana imbibed water during soaking, indicating that they were permeable and that the delayed germination was not due to a water impermeable barrier. However, the nuts imbibed water more slowly and had significantly less mass than seeds without the pericarp after 240 h of soaking. Thus, the pericarp is permeable;however,it inhibits the water uptake to some degree.
Corylus avellana seeds have been classified as recalcitrant because dormancy is induced by drying (Hong and Ellis 1996)and based on the ratio of the total seed dry mass for cotyledons and embryonic axis even thouhg they can withstand much greater dessication than is typical for recalcitrant seeds (Normah et al. 1994). From the result of the experiment on the viability of seeds after drying, C.avellana seeds clearly can withstand desiccation to low moisture contents. No viability was lost when seed moisture content was reduced to 8.71%; the germination percentage (regardless of CS period) of desiccated seeds did not differ significantly from those with 20.73% MC. This result indicates that C. avellana seeds do not satisfy the definition of recalcitrant seeds, since the seeds of the species maintain viability at low level of MC(8.71%),enough lower that the typical MC of seeds in this category.Similar results were obtained by Michalak et al. (2013) in a study on germination response of C. avellana seeds to dehydration and exposure to very low temperatures.They reported that C. avellana seeds tolerate strong desiccation (up to 2.7%) and, agreeing with Normah et al. (1994), should be classified as orthodox and not as recalcitrant. However,more studies are needed to determine whether C. avellana seeds should be classified as orthodox or suborthodox (an intermediate category).

Fig. 3 Effect of sowing date (filled diamond-15 September, filled square-2 November and filled triangle-17 December) on seedling emergence (percentage of sown nuts) in the following spring. Means with different letters differed significantly at p <0.05
A 2-month period of CS was needed for complete dormancy breaking and the highest germination (see Table 1)of true seeds.In the case of nuts,however,CS only was not very effective in breaking the dormancy; nuts cold-stratified up to 3 months had low GPs (up to 36%,Table 2).Thus, the pericarp, although permeable to water, is inhibitory to seed germination,and CS only(up to 3 months)is not sufficient to release dormancy induced by the pericarp.According to Bradbeer et al. (1978), inhibitors in the pericarp, testa and possibly mechanical resistance of the pericarp inhibit germination in freshly harvested nuts of C.avellana. The removal of pericarp and testa in freshly harvested C. avellana nuts results in a high germination rate (Jarvis 1975; Bradbeer et al. 1978). In the present study, removing only the pericarp of nuts stored for 5 months at 3-5 °C resulted in about half of the seeds to germinate (see Table 1). However, according to Jarvis(1975), embryo dormancy develops during dry storage of C. avellana nuts and he demonstrated that dry storage longer than 24 days induces full embryo dormancy, and consequently, seeds without the pericarp and testa do not germinate. The embryo dormancy was attributed by Jarvis(1975) and Bradbeer (1968) to the movement of inhibitors from the pericarp and testa into the cotyledons, and subsequently into the embryonic axis. Similarly, Frankland and Wareing (1966) stated that the ability of seeds after removal of pericarp and testa to germinate is lost after dry storage of C. avellana nuts for 8 weeks at 20 °C. Furthermore, Jarvis and Wilson (1977) stated that embryo dormancy in C. avellana seeds is induced after a short period of storage of nuts at laboratory temperature. In the present study, low temperature (3-5 °C) and relative high moisture content of seeds(20.73%)during the 5 months of storage might led to germination of a significant proportion of seeds after removing only the pericarp. Cold stratification treatment promoted germination, and 2 months was needed to maximize germination. This result is consistent with the studies of Bradbeer et al. (1978) and Jarvis and Wilson (1977) who used CS and gibberellic acid to overcome embryo dormancy in C. avellana seeds without the pericarp.
Before the preset study, there was no information on overcoming dormancy of C. avellana nuts. In germination tests using nuts, dormancy was entirely broken and the maximum germination (95%) achieved only when a 2-month WS preceded a 2-month CS. Warm stratification treatment may help to weaken the pericarp and make it more permeable to water, thus allowing the emergence of radicle and/or significantly reduce the concentration of substances which inhibit the germination. Possibly, a 1-month WS before the CS was not sufficient to release pericarp dormancy. According to Macdonald (2006) and Hartmann et al.(2011),the WS is required to overcome the dormancy imposed by a hard endocarp. The beneficial effect of warm stratification on seed germination of Crataegus monogyna species could be attributed to a decrease in pericarp mechanical resistance after this treatment (Persson et al. 2006). Furthermore, it is well known that during stratification, the balance between inhibitors and promoters in seeds changes. For example, the abscisic acid (ABA) concentration in the endocarp of Prunus campanulata seeds, is significantly reduced during warm stratification (Chen et al. 2007). Abscisic acid regulates seed dormancy and germination (Nambara et al. 2010).However, the levels of gibberellins (GAs, which promote seed germination) in embryo increase during cold stratification(Powell 1987;Chen et al.2007).Possibly,the length(up to 3 months) of the CS used alone in the present study increased the GAs, but did not sufficient decrease ABA,whereas ABA levels might be significantly reduced only after the WS is followed by CS.
The need for WS of nuts was also confirmed by the outdoor sowing results. Sowing nuts immediately after collection in early autumn exposed seeds to a sequence of autumn →winter temperatures in the presence of moisture, thus breaking dormancy and achieving maximum seedling emergence in the following spring. The moisture content of the substrate where the nuts were sown was regulated by watering.The early autumn sowing permitted exposure to warm temperatures for at least 2 months(Fig. 1) before winter temperatures, resulting in about 83.33% seed germination in the spring. Nuts sown later (2 November)received fewer days of warm temperatures,and germinated reached only 62.50%. According to Aygun et al. (2009), germination of C. colurna nuts sown in an outdoor nursery in November was 64%the next spring.On the other hand, Ninic-Todorovic (2000) suggested that C.colurna nuts should be sown in October rather than in September or November for the best results. In our study,C. avellana nuts sown on 17 December, with the exposure to cold only had the lowest germination in the spring(44.17%). This need for exposure to a period of warm temperatures before winter to maximize spring seedling emergence has been noted for many species with seeds enclosed by a hard pericarp (Pipinis et al. 2017).
Conclusions
In C.avellana,the pericarp is not a barrier to water uptake by the seed.The seeds tolerate desiccation(8.71%moisture content), and therefore, should not be classified as recalcitrant. Cold stratification at 3-5 °C for 2 months was needed to maximize germination of seeds after the pericarpremoval of nuts that had been stored for 5 months at 3-5 °C.For large quantities of nuts,removing the pericarp is not practical, so a 2-month warm stratification at 20/25 °C followed by a 2-month cold stratification is proposed as an effective pretreatment to enhance seed germination. Because the sowing date affected the spring emergence of seedlings from nuts sown outdoors, the nuts must be sown in the nursery immediately after collection in early autumn so that the natural sequence of temperature conditions will fulfill requirements for complete dormancy release and maximum germination and seedling emergence.
杂志排行

Journal of Forestry Research的其它文章
- Do increasing respiratory costs explain the decline with age of forest growth rate?
- At what carbon price forest cutting should stop
- Mapping the risk of winter storm damage using GIS-based fuzzy logic
- Comparison of seed morphology of two ginkgo cultivars
- De novo assembly of the seed transcriptome and search for potential EST-SSR markers for an endangered, economically important tree species: Elaeagnus mollis Diels
- Effect of artificially accelerated aging on the vigor of Metasequoia glyptostroboides seeds