Leaf area index estimated by direct, semi-direct, and indirect methods in European beech and sycamore maple stands
2020-05-19JakubPavelHaninecRadekPokorn
Jakub · Pavel Haninec · Radek Pokorný
Abstract Leaf area index (LAI) is one of the most important characteristics of forest stands that affects the fundamentals of tree physiological processes, biomass production, and mechanical stability. The LAI results obtained by the semi-direct and indirect methods (the needle technique and an LAI-2000 PCA)in three European beech(Fagus sylvatica L.)stands and one sycamore maple(Acer pseudoplatanus L.) stand were compared with LAI estimated by litter traps during the 2013 growing season.Seasonal LAI was estimated using an LAI-2000 PCA which showed similar trends among the stands and strongly corresponded to phenological phases of deciduous stands in Europe, with the fastest rate of leaf area increment occurring during the first month following bud break.During the growing season, maximum stand LAI value was on June 19th and reached 4.5-5.1, and 4.0 in the beech and maple stands, respectively. The needle technique significantly underestimated(p <0.05)direct LAI on average by 22.0%and 40.0%in the beech and maple stands,respectively.The LAI-2000 PCA insignificantly underestimated (p >.05)LAI on average by 15.1%and 5.8%in the beech and maple stands, respectively.All methods for LAI estimation at the stand level could be applicable in deciduous forest stands(beech, maple) with similar site and stand characteristics.However, calibration by direct method is necessary to obtain the required precision.
Keywords Leaf area index · Specific leaf area · Litter trap · Needle technique · LAI-2000 PCA
Introduction
Leaf area index(LAI)estimation is prominent in studies of terrestrial ecosystems because it is an important determinant of carbon, water, and energy exchange at the stand level (Norman 1992), and at landscape (Running et al.1989) and global scales (Wittwer 1983; Botkin 1986). It is defined as the total one-sided leaf area per ground surface area (Watson 1947; Marshall and Waring 1986; Chen and Black 1991; Lang et al. 1991).
The seasonal cycle of LAI determines many key aspects of forest canopy functioning (e.g., radiation extinction,rainfall interception, photosynthetic uptake, and evapotranspiration; Saigusa et al. 2002). Typically, the seasonal LAI pattern with rapid leaf development in the spring, a constant maximum LAI value in the summer, and an LAI decline in autumn has been observed in temperate deciduous stands (Gond et al. 1999; Mussche et al. 2001). The maximum LAI of European deciduous stands commonly occurs close to mid-July (Le Dantec et al. 2000; Mussche et al. 2001).
During the growing season, LAI can be determined by direct, semi-direct, or indirect methods. The beginning of the growing season is defined as the first day after 5 consecutive days where the average daily temperatures are higher than 5 °C (Ta>5 °C), and the end of the growing season is the first day after 5 successive days where the average daily temperatures are Ta<5 °C(Krupková et al.2017). The direct method involves harvesting and litter collection (Bréda 2003). Given that leaf harvesting is extremely labour-intensive, time-consuming, and requires many replicates to reduce sampling errors, especially within tall, heterogenous canopies such as those in deciduous forest stands(Thimonier et al.2010),researchers have developed less destructive methods such as litter traps(Bréda 2003) and the semi-direct needle technique(Dufrêne and Bréda 1995; Bréda 2003). The litter trap technique is simple,however it is based on the assumption that the traps collect a random sample of falling leaves,that collection is made every second week to avoid losses and decomposition during leaf senescence (Bréda 2003), and that the contents of the traps represent the average amount of leaves falling in the stand (Eriksson et al. 2005). In this technique, the litter is dried to a constant weight and used to calculate litter dry mass as g m-2. This dry mass is converted into leaf area by multiplying the biomass by the specific leaf area value(SLA;m2g-1)or leaf dry mass perarea(LMA,g m-2)as inverse of SLA(čermák 1998).The LAI is obtained throughout complete leaf senescence(Bréda 2003).This litter trap method is highly accurate and has been successfully applied in deciduous forests in Tennessee (Chason et al. 1991), and may be used to determine the LAI of other deciduous forests(Marshall and Waring 1986; Neumann et al. 1989).
LAI may be also assessed by the semi-direct needle technique which determines leaf area from the inclined point quadrate method (Goodall 1952; Warren Wilson 1959, 1960, 1963). A thin, sharp needle is plunged vertically into the litter on the ground.With a vertical probe and horizontal leaves, the number of leaves collected on the needle is related to the contact number and equals the LAI.The needle technique requires an intensive sampling from 100 to 300 points per stand per probe to quantify an average contact number and to correctly derive LAI(Nizinski and Saugier 1988; Dufrêne and Bréda 1995;Bréda 2003). These two methods are based on the assumption that total leaf number and area on the tree is the same as on the soil surface after leaf senescence (Dufrêne and Bréda 1995).
Indirect methods are based on more easily measurable parameters to derive LAI (Fassnacht et al. 1994; Gower et al. 1999). These are: (1) canopy gap fraction and/or (2)radiation transmittance (Gower and Norman 1991). These indirect methods relate one-sided leaf area to the radiation environment below the canopy through the inversion of an appropriate radiative transfer model(Anderson 1971;Ross 1981; Norman and Campbell 1989). In the most widely applied indirect techniques, LAI was inferred from the inversion of the model describing the attenuation of radiation passing through the canopy and/or using gap-fraction data evaluated over a range of zenith angles (Thimonier et al. 2010). One of the most commonly used instruments for estimating gap fractions is the LAI-2000 PCA (LICOR®, NE, USA) because its sensors assess the canopy gap fraction from several different zenithal angles simultaneously (Küßner and Mosandl 2000; Thimonier et al.2010) within the blue light wavelength interval of photosynthetic active radiation (PAR <490 nm; Pokorný and Marek 2000; Mason et al. 2012). In this blue wavelength,the leaves reflect and transmit relatively negligible amounts of radiation.The foliage appears black against the sky(LICOR 1991). The LAI-2000 PCA can be used during an entire growing season (Mussche et al. 2001).
A number of studies have compared LAI values found by the direct method with those indirectly estimated(Sampson and Allen 1995; Eschenbach and Kappen 1996;Olivas et al. 2013; Behera et al. 2015), and/or indirect methods compared to each other have been reported(Garrigues et al. 2008; Eckrich et al. 2013). There were typically strong correlations between LAI assessed using an LAI-2000 PCA and litter traps (Behera et al. 2015); in many cases the LAI-2000 PCA led to underestimating stand LAI (Dufrêne and Bréda 1995; Cutini et al. 1998;Mussche et al.2001).However,the comparisons of directly estimated LAI and the semi-direct needle method have been absent in the literature over the last decade (Dufrêne and Bréda 1995; Bréda 2003).
The objectives of this study are: (1) to observe accurate seasonal LAI dynamics using an LAI-2000 PCA in chosen deciduous stands; (2) to estimate the maximum stand LAI within the growing season indirectly using an LAI-2000 PCA; and, (3) to compare LAI values obtained by three distinct methods of estimation (direct, semi-direct, and indirect) at the stand level in selected beech and maple stands.
Materials and methods
Study site
All measurements were conducted on four research plots,with three plots located at the Training Forest Enterprise Masaryk Forest (TFE MF) Křtiny and one plot situated in the vicinity of the Long-term Experiment Station Rájec-Nĕmčice (Table 1; Fig. 1).
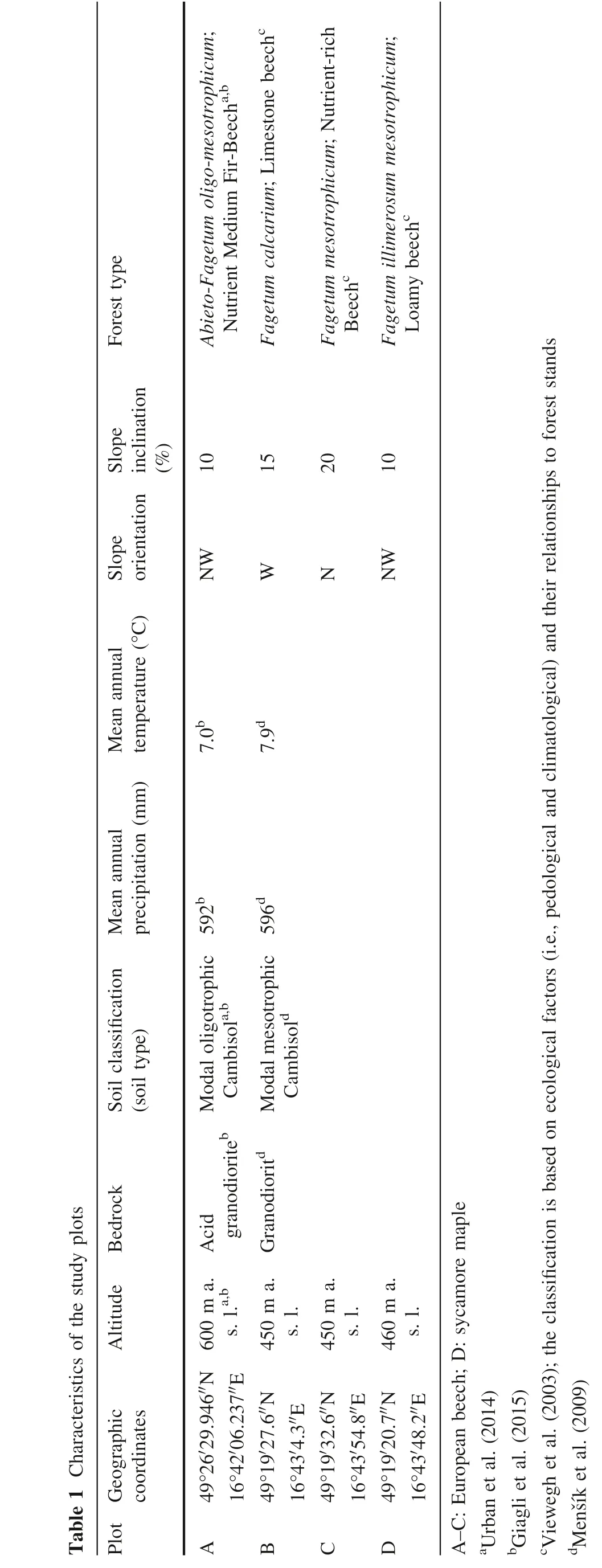
Table 1 Characteristics of the study plots Forest type Slope inclination(%)Slope orientation Mean annual temperature (°C)Mean annual precipitation (mm)Soilclassification(soil type)BedrockAltitude Plot Geographic coordinates Abieto-Fagetum oligo-mesotrophicum;Nutrient Medium Fir-Beecha,b Fagetum calcarium; Limestone beechc Fagetum mesotrophicum; Nutrient-rich Beechc Fagetum illimerosum mesotrophicum;Loamy beechc 10152010 NW W N NW 7.0b 7.9d 592b 596d Acidgranodioriteb Modal oligotrophic Cambisola,b Modal mesotrophic Cambisold Granodioritd 600 m a.s. l.a,b 450 m a.s. l.450 m a.s. l.460 m a.s. l.49°26′29.946′′N 16°42′06.237′′E 49°19′27.6′′N 16°43′4.3′′E 49°19′32.6′′N 16°43′54.8′′E 49°19′20.7′′N 16°43′48.2′′E A B C D A-C: European beech; D: sycamore maple aUrban et al. (2014)bGiagli et al. (2015)cViewegh et al. (2003); the classification is based on ecological factors (i.e., pedological and climatological) and their relationships toforest stands dMenŠík et al. (2009)

Fig. 1 Location of the research plots. A, B, C: European beech; D: sycamore maple
Square plots with 20 m sides were established using the Field-Map®technology (IFER, Czech Republic). Tree heights were determined using an electronic laser hypsometer (Vertex®Laser VL5; Haglöf, Sweden) with an accuracy of 0.1 m, and diameters at breast height (DBH)were measured with a calliper with a precision of 0.1 cm.Dendrometric and structural characteristics of the plots are listed inTable 2.
LAI estimation
Stand LAI was estimated using three distinct methodological approaches (i.e., directly, semi-directly, and indirectly by litter traps, the needle technique, and an LAI-2000 PCA, respectively).
LAI determined with litter traps was used as the reference when comparing the other two (Dufrêne and Bréda 1995; Bréda 2003; Jonckheere et al. 2004). The precision of the direct method depended on the number of traps,their distribution below the canopy, and the speed and direction of the wind.Therefore,15 litter traps with a capture area of 0.25 m2(Dufrêne and Bréda 1995;Cutini et al.1998)were randomly distributed throughout the plots 1 m above the ground (McShane et al. 1983; Morrison 1991; Cutini et al.1998).The direct LAI estimation was applied every second week when 10% leaf colour was observed, i.e., from mid-September (see Table 1 and Fig. 2B in Urban et al. 2014),to avoid leaf losses by wind and decomposition (Bréda 2003). For each plot, litterfall collected by traps was removed of other materials (branches, seeds) and ovendried for 48 h at 60 °C to reach a constant weight(Garnier et al. 2001; Vile et al. 2005; Homolová et al. 2013). To obtain the stand LAI, the total dry leaf mass per unit of ground area was multiplied by the weighted mean SLA(i.e., fresh leaf area per unit of leaf dry mass). The SLA was assessed as a sub-sample represented by the leaves of one trap at each collection period in each plot due to the considerable differences in SLAs collected by various traps on the same plot. The exact area of the leaves wasdetermined using the ImageJ®software (Wayne Rasband,MD, USA).

Table 2 Dendrometric characteristics (Mean ± SD) of the stands
The semi-direct needle technique was carried out and is based on an inclined point quadrat(cf.introduction part).It was carried out once immediately after leaf senescence(i.e., the end of October). On each plot, 100 sampling points were randomly selected and needle penetrations performed at a 57° angle (according to čermák, personal communication) through the layer of freshly fallen leaves on the ground surface (Dufrêne and Bréda 1995; Bréda 2003).The technique used a sharp metallic needle 1.5 mm in diameter,and the average number of leaves pierced with each stab was equivalent to the LAI (Nizinski and Saugier 1988).
LAI was likewise estimated using an LAI-2000 PCA(LI-COR®,NE,USA) at 27 regularly spaced points within the permanent transects (Chen and Cihlar 1995a, b; Battaglia et al. 1998; Majasalmi et al. 2012). These 27 points were divided into two perpendicular transects with 13 measurement points located within each transect (i.e., 26 together)and with one as the 27th and central point.These permanent transects were established to monitor seasonal LAI to avoid any errors due to different views of the sensors on the plots.The LAI was obtained monthly in the same unit using an identical procedure throughout the growing season from April until leaf senescence in November 2013. For more detailed seasonal LAI observations and the date of maximum LAI,measurements were repeated every 2 weeks on plot A. LAI was taken solely under diffuse radiation during windless conditions (Cˇater et al. 2013). The first and last measurements were always taken in a sufficiently large open area. During all measurements, the LAI-2000 PCA sensor was held 1.3 m above the ground (Holst et al. 2004) and a 180° view restrictor used to exclude the operator from the viewing area. Because Kucharik et al. (1998) noted that LAI is substantially affected by the presence of stems (stem area index; SAI) and branches (branch area index; BAI), the first and last measurements were taken to determine the woody area index (WAI; WAI = SAI + BAI) before bud break and after leaf senescence (Chason et al. 1991;Dufrêne and Bréda 1995;Cutini et al.1998).To obtain the true stand LAI, the mean WAI was subtracted from plant area index (PAI; LAI = PAI - WAI). The resulting dataset was processed using FV2200 software (LI-COR®, NE,USA). The innermost sensor rings with a maximum zenith angle of 43° were included (Küßner and Mosandl 2000;Pokorný and Marek 2000; Pokorný and Stojnič2012) to avoid edge effects in an open area (Dufrêne and Bréda 1995), and the default program setting was selected for all other parameters.
Statistical data analyses
Statistical analyses were performed using STATISTICA®software, version 10.0 (StatSoft Inc., OK, USA) and SigmaPlot®software, version 13.0 (Systat Software Inc., CA,USA) using the confidence level of 0.95. The Shapiro-Wilk’s test was applied for testing the normality of data distribution. If normal data distribution was confirmed,parametric tests (t test, ANOVA) were performed. Data with abnormal distributions were processed by the nonparametric Mann-Whitney test. A variance analysis was carried out for detecting the occurrence of significant differences among the tested groups.The Tukey post-hoc test was used tofind significant differences among individual groups.A t test was performed when only two groups were compared (e.g., between measurement approaches).Finally, descriptive statistics for each dataset were calculated.
Results
In April,when the buds had not broken,the LAI was zero.At this time,WAI was simply estimated;its mean seasonal values for plots A, B, C, and D were 1.33, 0.26, 0.99, and 0.38, respectively (Fig. 2). The most progressive LAI development was during the period from bud break in April until the start of May (part 1, Fig. 2). From May until the end of June (part 2, Fig. 2), the leaves continued to develop, however with less intensity compared to part 1.LAI dropped by 0.46 from the second half of June until the end of July on plot B. On plot A, seasonal LAI measurements were taken at shorter time intervals. Therefore, the stagnation of the LAI was more obvious during the summer months (part 3, Fig. 2). At the end of September, leaves started tofall in all stands, illustrated by the decline in the LAI curve (part 4, Fig. 2).
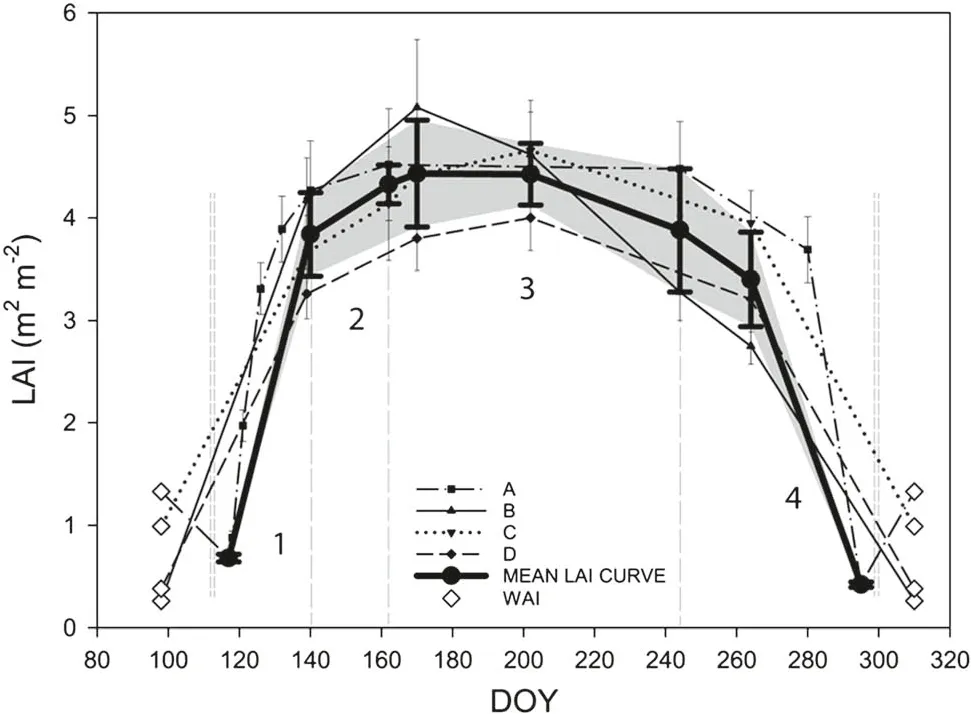
Fig. 2 Seasonal LAI dynamics during the 2013 growing season. LAI leaf area index;WAI woody area index;A-C:beech research plots;D:maple plot; DOY day of the year. Empty diamonds signify average seasonal WAI subtracted from the plant area index (PAI) to obtain true LAI (LAI = PAI - WAI). Period 3 appears to be the most suitable phase to compare the LAI of deciduous stands during the growing season. The whiskers display standard deviations of LAI estimation and the grey area signifies the confidence interval of the mean LAI curve
Average LAI values of all stands during the 2013 growing season are exhibited in Fig. 3. The highest values were obtained by the litter traps on all plots except A. The needle technique gave the highest average LAI value in plot A. The differences between LAI values estimated by the direct method(litter traps)and an LAI-2000 PCA were insignificant(p >.05;Fig. 3,left).In plots B,C,and D,the needle method significantly underestimated the LAI obtained from the litter traps. Conversely, in plot A, the needle technique overestimated the LAI obtained by the direct method,but insignificantly(p = 0.1;Fig. 3,middle).The differences among the LAI values estimated by an LAI-2000 PCA and the needle technique were statistically significant (p <0.001) in all plots (Fig. 3, right).
Figure 4 shows the deviations of underestimation or overestimation of LAI using an LAI-2000 PCA and the needle technique, with both compared to the reference(litter traps). The LAI underestimations between the reference and the LAI by an LAI-2000 PCA on plots A,B,C,and D were 15.3%,11.0%,18.9%,and 5.8%,respectively.The average deflection of the LAI values on the beech plots and all probed plots were 15.1% and 12.7%, respectively.On plots B, C, and D, the needle method underestimated the direct method by 41.0%, 38.0%, and 40.0%, respectively. A 13.0% overestimation of LAI by the needle technique versus the direct method was observed on plot A.Mean deviations of the LAI values in beech and in all plots regardless of tree species were 39.7% and 26.5%,respectively.

Fig. 4 Mean deflection of LAI values estimated by the needle technique and the LAI-2000 PCA from the reference(litter traps).AC: beech research plots; D: maple plot; ALL-mean deviation of all plots regardless of tree species. The whiskers show standard deviations
Discussion
Seasonal LAI development revealed a typical pattern during the growing season,reflecting the leaf phenology of the species under their site conditions. In the beech vegetation zone in the Czech Republic(i.e.,at elevations from 550 to 600 m a.s.l., with a mean annual temperature range of 6.5-7.5 °C, an annual amount of precipitation varying between 690 and 800 mm, and a growing season of 140-150 days), the beginning of bud flushing and tree growth usually starts at the end of April and lasts until the end of September when leaves begin falling. Mean daily temperature is an average from three air temperature readings taken at 7 a.m., 2 p.m., and 9 p.m. CET (Central European Time) at 2 m above the ground surface by a covered sensor (www.chmi.cz). Bréda (2003) recorded an almost identical pattern of LAI development in beech experimental plots in northeastern France with similar site conditions. The LAI pattern within the growing season on five beech plots is described by Bequet (2011) who alsonoted a similar LAI seasonal pattern as in this study. He divided the LAI seasonal curve into three phases whereas we divided the growing season intofour which brings greater detail to leaf phenology and LAI changes. On plot A, where the LAI seasonal pattern was observed in short time steps,a moderate stagnation of LAI was detected only in the summer. This corresponds to the results from the study by Bréda (2003). Therefore, the best period for comparing LAI estimates by the distinct methods occurs during this time (days 140-240, Fig. 2). On plot B, seasonal LAI slightly increased within the June-July period when the majority of Central European deciduous tree species commonly reach their seasonal maximum LAI value (e.g., Bréda and Granier 1996; Le Dantec et al.2000), and then stagnate for a period. This was also reported by Bequet (2011), when a small variation of LAI may be explained by either a measurement error and/or a pest outbreak whose attack reduces leaves (Zvereva et al.2012). Plot B is exposed to direct sunlight for the longest period of time; this could result in a stronger short-term drought within the summer months, causing partial leaf senescence(Coder 1999).Mussche et al.(2001)observed a similar seasonal LAI pattern in a mixed oak-beech stand and a pure ash stand in Belgium. The spring development of LAI relating to the results of this research and to those of Bréda (2003) indicate that maximum LAI is commonly reached at the end of June. Mussche et al. (2001) reported stagnation from the beginning of September, which is similar to plot A. Conversely, their LAI monitoring also included a measurement showing unexpectedly increasing LAI from the beginning of June to July,then dropping and stagnating until September, and decreasing similarly as on plot B, which could have been caused by a drought during this time. During an extreme summer drought leading to defoliation, reflushing can subsequently occur (Filewod and Thomas 2014). Commonly, secondary flushing exists when Lammas shoots/sprouts arise in mid- or the end of June.

Fig. 3 Comparison of the statistically significant differences amongbeech research plots; D: maple plot; p <0.05 (*); p <0.001 (**);average LAI values estimated by the direct (litter traps), semi-directp >.05 (ns). The whiskers show standard deviations(the needle technique), and indirect (LAI-2000 PCA) methods; A-C:
On the beech plots (A, B, and C), the maximum LAI values estimated by litter traps, the needle technique, and an LAI-2000 PCA ranged between 5.2-5.6, 3.4-6.0, and 4.5-5.1,respectively.On the maple plot(D),the maximum LAI values obtained by litter traps, the needle technique,and an LAI-2000 PCA reached 4.2, 2.5, and 4.0, respectively. The mean seasonal maximum LAI value of pure beech stands estimated by Michelot et al. (2012) was 5.1.The reported values from litter traps and an LAI-2000 PCA are fully comparable, except for those estimated by the needle method; this method shows highly varying and often very low values.Le Dantec et al.(2000)estimated the maximum LAI value of 5.0 using an LAI-2000 PCA in a mature beech stand with a breast height basal area of 31.9 m2ha-1in central France. Their seasonal maximum LAI value agreed well with the LAI on plots A and C,which reached 4.5 and 4.7 with basal areas of 38.8 and 40.9 m2ha-1, respectively. In Italy, beech stands reached maximum LAI of 6.0(from litter traps)and 4.9(LAI-2000 PCA)(Cutini et al.1998).Chianucci et al.(2016)estimated stand LAI values of 6.1 and 5.0 using an LAI-2000 PCA in control and thinned beech stands, respectively. These results from the LAI-2000 PCA confirmed the measurement outputs for all beech plots in this study.Pokorný and OpluŠtilová (1999) noted that maximum LAI of deciduous stands in the Czech Republic estimated by an LAI-2000 PCA varied between 4.0 and 8.0. In beech stands with similar site and weather conditions, these outputs and the results of this paper agree with studies by Bartelink(1997),Mussche et al. (2001) and Bequet (2011), who reported average LAI values of 4.9, 4.0, and 4.2 respectively. Jurik et al. (1985) assessed the maximum LAI by direct harvesting in young aspen-oak-maple stands with similar basal area (6.5 cf. 6.9 in this study). They published a lower stand maximum LAI value of 2.3 than we have noted.The discrepancy may be due to a high aspen and oak mixture,amplified by a lower basal area. It appears as if LAI between 5.0 and 6.0 indicates healthy and productive beech stands, however a test of LAI efficiency in above-ground biomass production should be performed (see Pokornýet al. 2008 for Norway spruce).
The LAI-2000 PCA method generally underestimated the LAI found from litter traps by 15.1% and 5.8% in the beech plots(A,B,C)and the maple plot(D),respectively.The most accurate method for LAI estimation is the direct method,which is based on harvest.Therefore,the litter trap method taken as an accurate standard. A possible inaccuracy of LAI estimation using litter traps may occur due to wind, the amount and distribution of the traps within the stand regardless of canopy cover and stand structure, and the size of the stand area (Finotti et al. 2003; Yang et al.2017). Cutini et al. (1998) showed that the average underestimated value of indirect methods compared to litter traps was 29.6% in beech stands. Bréda(2003) noted that measuring LAI using an LAI-2000 PCA underestimates the litter trap values by an average of 32.0%,almost double the value found in this study.Mussche et al.(2001)compared direct and indirect LAI estimations using litter traps and an LAI-2000 PCA in beech-oak stands dominated by beech.The LAI-2000 PCA underestimated the litter trap values by approximately 18.5%. Thus their reported deviations among direct and indirect methods were similar to this study (cf. Fig. 4). The deviations between LAI values could be caused by factors influencing LAI estimation using an LAI-2000 PCA (e.g., non-random leaf area distribution,the angle of the sensor view versus the tree crown base, foliage clumping within the shoots, the number and distribution of sample points owing to canopy closure and/or the position of the stems in the stands). There may also be a possible error affecting the litter traps evaluation(e.g.,trap distribution; unhomogenised sample of the SLA; the capturing area of the traps; and/or the speed and direction of the wind). Dufrêne and Bréda (1995) reported systematically lower LAI values of 6.0-37.0% by needle technique compared to the direct one in temperate deciduous stands. However, considering only the pure beech stand investigated by them, the needle technique underestimated the direct method by approximately 18.0%. In this study,underestimations of 41.0%, 38.0%, and 40.0% were reported for plots B, C, and D, respectively. Conversely,the needle technique overestimated the value of the direct method by 13.0% on plot A. On all plots, the detected twofold deflection could be influenced by the number and distribution of the sample points, leaf decomposition, and/or the possible effect of wind below the canopy (i.e., an uneven litterfall could accumulate close to trunk base). In addition, the needle technique could be dependent on needle diameter and angle of penetration through the leaffall (Dufrêne and Bréda 1995). On plot A, the overestimation cf. the direct method could also have been caused by the angles among the newly fallen leaves and the horizontal ground surface and/or by not maintaining the same angle of penetration (i.e., 57°).
Conclusions
The direct, semi-direct, and indirect methods were suitable for LAI estimation of the beech and maple stands.The results indicate a strong relationship between LAI values using an LAI-2000 PCA and litter traps, with insignificant(p >.05) differences between them. The LAI-2000 PCA underestimation of litter trap values were 15.0% and 6.0%in beech and maple stands, respectively. There were significant differences between litter trap and needle technique values within three of the four forest plots(p <0.05). The needle technique underestimation compared to the direct reference method varied from 22.0 to 40.0%.However,the technique appears to be practical and less time-consuming and laborious than litter traps. The results vary considerably and LAI estimation accuracy depends more on the number of probe penetrations as leaves accumulate in some micro-places or are blown away than on the angle of probe insertion.From a practical point,the indirect methods of LAI estimation appear better even though inaccuracy may reach approximately 15.0%.Obtaining seasonal maximum LAI from litter traps is possible only after summarizing all fallen leaves during the growing season. Therefore, calibration with the direct method by assessing a site-specific correction factor is necessary. The seasonal LAI pattern, as well as the maximum stand LAI during the growing season (DOY 170), fit well with the phenological phases for both beech and maple species. Based on these results, we suggest that the LAI-2000 PCA is applicable in deciduous forest stands with similar site and stand characteristics.
AcknowledgementsThe English language was kindly proofread by the BEST translation agency.Special thanks to Dr.Pavel Bednářand three anonymous reviewers for their helpful and valuable comments on improving this paper.
杂志排行

Journal of Forestry Research的其它文章
- Do increasing respiratory costs explain the decline with age of forest growth rate?
- At what carbon price forest cutting should stop
- Mapping the risk of winter storm damage using GIS-based fuzzy logic
- Effects of seed moisture content, stratification and sowing date on the germination of Corylus avellana seeds
- Comparison of seed morphology of two ginkgo cultivars
- De novo assembly of the seed transcriptome and search for potential EST-SSR markers for an endangered, economically important tree species: Elaeagnus mollis Diels