Effects of an ectomycorrhizal fungus on the growth and physiology of Pinus sylvestrisvar. mongolica seedlings subjected to saline-alkali stress
2020-05-19DachuanYinSaiyaremuHalifuRuiqingSongJinyuQiXunDengJifengDeng
Dachuan Yin · Saiyaremu Halifu · Ruiqing Song · Jinyu Qi · Xun Deng ·Jifeng Deng
Abstract This research investigates the mechanism of increased salinity tolerance of ectomycorrhizal fungiinoculated P. sylvestrisvar. mongolica to provide a theoretical basis for the application of the fungus in saline soils.Growth effects due to inoculation of seedlings with Suillus luteus (a symbiotic ectomycorrhizal fungus), were determined in four kinds of saline-alkalisoils. Growth and physiological indicators, including photosynthetic characteristics, plant height, biomass, photosynthetic pigments,catalase (CAT) and superoxide dismutase (SOD) enzyme levels,and malondialdehyde(MDA),an organic marker for oxidative stress, and soluble protein levels were determined. Mycorrhizal colonization rate decreased with increasing saline-alkalinity and growth of inoculated seedlings was significantly enhanced. Biomass and chlorophyll contents also increased significantly. SOD and CAT activities were higher than in non-inoculated seedlings. However, MDA content decreased in inoculated seedlings. Soluble protein content did not increase significantly. Inoculation with a symbiotic ectomycorrhizal fungus could enhance the saline-alkali tolerance of P. sylvestrisvar. mongolica. Growth and physiological performance of inoculated seedlings were significantly better than that of uninoculated seedlings. The results indicate that inoculated P. sylvestrisvar. mongolica seedlings may be useful in the improvement of saline-alkali lands.
Keywords Ectomycorrhizal fungi · Saline-alkali stress ·Pinus sylvestrisvar. mongolica · Physiological and biochemical mechanisms
Introduction
Environment degradation is a major issue impacting human development and survival in the twenty-first century(Yang et al.2005;Ravi et al.2010),particularly in arid and semiarid areas(Stocker et al.2013).With rapid global warming,regional temperatures will rise, droughts will intensify and soil salinization will increase, resulting in a profound impact on the health of forest ecosystems (Gauthier et al.2015; Lugo 2015; Sugden et al. 2015; Trumbore et al.2015). Soil salinization is a growing environmental problem, particularly in the arid and semi-arid areas of northeastern China which has the largest saline-alkaline areas in the country. Alkaline meadows cover >70% of the land and this is increasing(Kawanabe and Zhu 1991).The soils are mainly affected by high levels of NaHCO3and Na2SO4salts. The high pH values cause saline and alkali stresses that damage plants,inhibiting ion uptake and disrupting the ionic balance of plant cells, thereby inhibiting growth and development (Yang et al. 2011).
Pinus sylvestrisvar. mongolica was introduced in the 1950s and is now one of the main afforestation species in northeast China.The trees grow rapidly,are highly tolerant to arid environments, and play important roles in local environments.
With the increase in salinization, P. sylvestrisvar.mongolica is under stress in northeastern China(Song et al.2015). This is manifest in the yellowing of needles and poor growth, leading to the decline of the tree (Zeng et al.1996;Wu et al.2004).Therefore,it is important to improve the survival rate of P. sylvestrisvar. mongolica in saline soils.One possibility of achieving this may be to introduce ectomycorrhizal fungi (ECMF) which are associated with the roots of Pinus species.This may stimulate plant growth and contribute to enhanced tolerance to abiotic stress(Abbaspour et al. 2012).
Ectomycorrhizal fungi can improve plant growth by increasing the absorption of mineral nutrients and water and increasing resistance to pathogens and environmental stresses (Shaw et al. 1995; Baar and Stanton 2000; Vinale et al. 2004; Mucha et al. 2006; Edda et al. 2010; Sharma et al. 2010). Inoculation with ECMF can enhance the ability of plants to resist salt stress by accumulating compatible solutes (Yin et al. 2017) and to adjust osmotic and ion balances(Yin et al.2014).However,the contribution of ECMF to growth and the physiological index of P. sylvestrisvar. mongolica under salt stress, particularly alkali stress, has rarely been reported.
Materials and methods
Organisms and experimental design
The study used a randomized block design with: (1)mycorrhizal inoculation(Suillus luteus(L.)Roussel and an uninoculated control); and (2) stress treatments-saline (10 and 20 mM Na2SO4)and alkali(10 and 20 mM NaHCO3).There were three pots in each treatment,each contained 15 seedlings.
Suillus luteus is the type species of the genus Suillus and was isolated from Horqin Sandy Land from a basidiome(sporocarp of the fungus) in a P. sylvestrisvar. mongolica plantation in Zhanggutai Liaoning Province, northeast China. The fungal spores were grown on MMN agar medium at pH 6.6(Yin et al. 2017).
Pine seeds,alsofrom Zhanggutai Liaoning Province,were surface sterilized with potassium permanganate (0.5%, v/v)for 30 min, washed three times with distilled water, and germinated on sterile,moistened gauze at 25 °C for 5 days.The germinants were transferred to plastic pots filled with vermiculite/soil/sand(1:2:1,v/v/v),which had been sterilized in a high-temperature autoclave for 2 h at 121 °C.Basic soil parameters were nitrogen (N) 520 mg kg-1, phosphorus(P)8.8 mg kg-1and potassium(K)76 mg kg-1,pH 7.3.
The pots were kept under greenhouse conditions,watered every 2 days for 1 month, and the seedlings were then inoculated with the fungus.
Plant culture and stress treatments
Each pot was treated with 100 mL of the inoculant or 100 mL of sterilized inoculant as the non-inoculated treatment.The inoculants were added 2 cm below the level of the seeds 90 days after sowing. Saline-alkali stress was created by adding the solutions described previously. The control seedlings were irrigated with distilled water. All treatments were arranged randomly and the seedlings harvested 10 days after treatment.
Growth index and mycorrhizal colonization rate
The roots were washed to remove soil. A total of 30 seedlings per treatment were randomly selected and height and biomass measured. Once fresh weight had been determined, the seedlings were oven- dried at 85 °C for 5 h. Twenty root sections were cut into 1-cm pieces.Mycorrhizal colonization rates were observed under a microscope.
The colonization rate (%) = mycorrhizal seedlings/total number of seedlings × 100.
Measurement of photosynthetic pigments
This followed the method of William and Paul (1985).Fresh needle tissue (0.5 g) was ground in 10 mL acetone and placed in a centrifuge at 10,000 rpm for 5 min.Absorbance was recorded at 663 and 645 nm. These experiments were repeated three times.
Malondialdehyde and soluble protein analysis
Malondialdehyde (MDA) levels were measured as described by Wasowicz et al. (1993). A 0.2 g amount of fresh needle tissue was placed in a test tube containing 1 mL distilled water.After adding 1 mL of 29 m mol L-1acetic acid, samples were placed in a water bath and heated for 1 h at 95-100 °C. After cooling under running water,25 μL 5 mol L-1HCl was added and the reaction was stopped by agitation for 5 min with 3.5 mL of n-butanol.After centrifuging at 10,000 rpm for 5 min, the separated butanol phase was removed and the fluorescence measured in a spectrofluorometer (Schimadzu-RF-5000, Kyoto,Japan) using 525 nm for excitation and 547 nm for emission.
Soluble protein levels were determined according to Christos et al. (2008). One hundred mg of Coomassie Brilliant Blue (CBB) G-250, 100 mg L-1reagent was added to 50 mL of 95%ethanol and mixed until dissolved.One hundred mL of 15 mol L-1H3PO4was added to 500 mL of distilled H2O and mixed well. A 0.5 g amount of needles were ground in 10 mL of distilled water and placed in a centrifuge at 10,000 rpm for 5 min. A one mL supernatant was added to 5 mL CBB reagent and mixed well. The absorbance was recorded at 595 nm.
Analysis of catalase and superoxide dismutase
The catalase (CAT) enzyme was extracted using 0.1 g of needles in 10 mL of 50 m mol L-1phosphoric acid buffer solution at pH 7.0. The extract was centrifuged at 10,000 rpm for 20 min at 4 °C and CAT activity analyzed.Superoxide dismutase (SOD) enzyme was extracted using 0.1 g of needle tissue in 10 mL of 50 m mol L-1phosphoric acid buffer solution at pH 7.8. The extract was centrifuged at 10,000 rpm for 20 min at 4 °C, and total SOD analyzed(Zhang et al.2010).All antioxidant enzyme activities were calculated following Song et al. (2005).
Statistical analysis
SPSS 13.0 software was used for the one-way analysis of variance (ANOVA) and two-way ANOVA testing at P = 0.05, P = 0.01, P = 0.001 of differences. The figures were drawn using Origin 8.5.
Results
Root colonization and growth
Mycelium colonization was not observed in roots of uninoculated seedlings. Inoculated seedlings displayed various degrees of colonization at different levels of saline-alkali stress (Fig. 1a, b). The colonization rate of mycorrhiza decreased with increasing degree of stress,and there were significant differences across all treatments.The maximum colonization rates in distilled water were 73%under saline and 75%under alkali stress.Colonization rate decreased with increasing salinity and alkalinity but the extent of reduction under alkali stress was higher than under saline stress.
Saline-alkali stress severely affects the growth of seedlings (Fig. 2a-f; Table 1). Inoculation with ECMF promoted the growth of the seedlings under saline-alkali stress. Height and biomass were significantly higher than for uninoculated seedlings (P ≤0.05) when [the concentration of saline or alkali]>0 mM. Heights of the inoculated seedlings were 18%, 13% and 7% higher than uninoculated seedlings (Fig. 2a). There were significant differences(P ≤0.05)across all treatments.The results of alkali stress are similar to those of salt stress with respect to height.In addition,seedling height was less with increasing saline-alkali stress. Seedlings wilted with the degree of stress but did not die.As height and biomass accumulation are correlated, similar results were found for biomass(Fig. 2c-f).
Photosynthetic pigments
Photosynthetic pigment content significantly decreased with increasing saline stress (Fig. 3a-d;Table 2). This is more pronounced when the concentrations of saline-alkali ≥10 mM (P ≤0.05). However, pigment content of+ ECMF seedlings decreased less than that of uninoculated seedlings with increasing saline-alkali concentrations. For chlorophyll a, there were no significant differences between inoculated and uninoculated seedlings at 0 mM saline and alkali concentrations (Fig. 3a, b). In contrast, for chlorophyll b, there were significant differences between the + ECMF and -ECMF treatments(P ≤0.05) when saline and alkali concentrations were 0 mM (Fig. 3c, d). Maximum photosynthetic pigment levels were obtained in distilled water in either saline or alkali stress, i.e., 0 mM concentration. This indicates thatthe photosynthetic characteristics of P. sylvestrisvar.mongolica seedlings significantly increased under salinealkali stress.
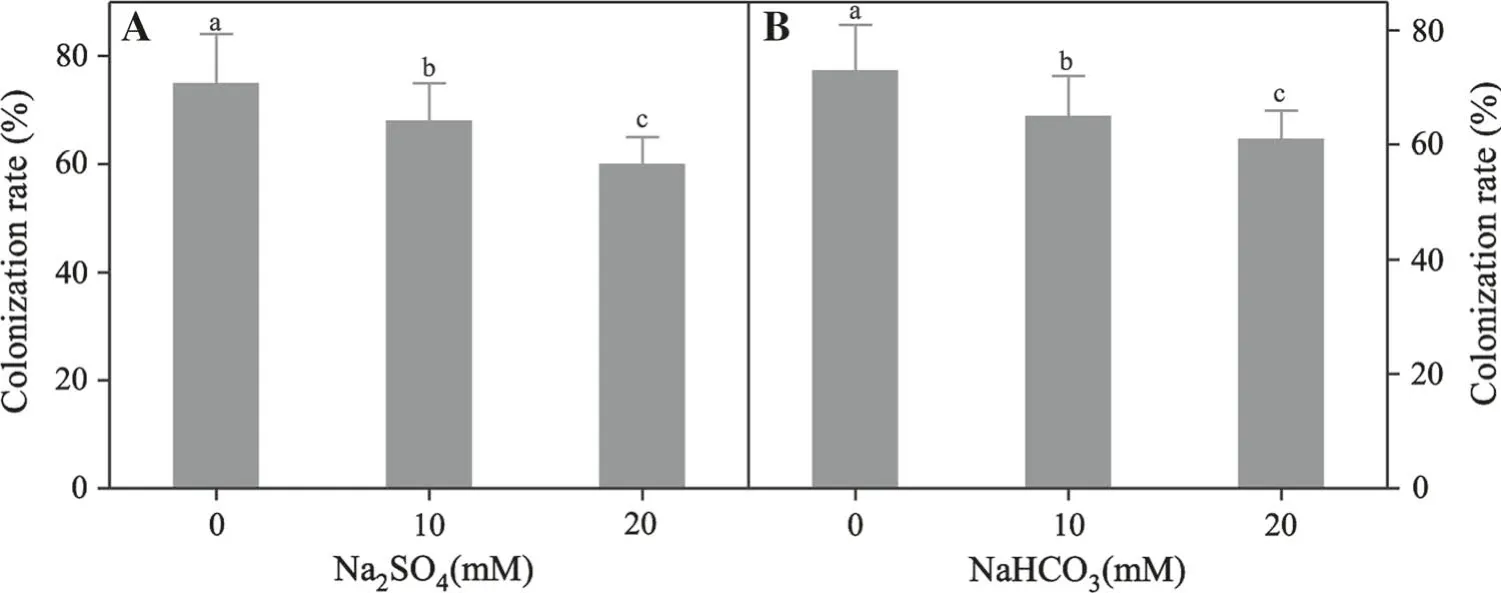
Fig. 1 Colonization rates under the different treatments(mean ± SD, n = 20) as affected by ECMF inoculation and under saline-alkali stress.Different letters are significantly different for each treatment(P ≤0.05)

Fig. 2 Growth indices(mean ± SD, n = 30) as affected by ECMF inoculation and under saline-alkali stress.Values with different letters are significantly different for each treatment (P ≤0.05)

Table 1 Results of two-way ANOVA on growth
Catalase and superoxide dismutase
The influence of ECMF inoculation on antioxidant enzymes was evaluated by determining CAT and SOD activity (Fig. 4a-d; Table 3). Inoculated seedlings under no-stress conditions displayed higher CAT and SOD activities than in other treatments. There were significant differences(P ≤0.05)in CAT activity between inoculated and uninoculated seedlings under saline-alkali stress. The greater the degree of stress, the greater the differences in CAT activity. In inoculated seedlings, CAT activity was 25% and 13% higher than the uninoculated seedlings at 20 mM saline and alkali treatments, respectively (Fig. 4a,b). Unlike CAT activity, there were no significant differences (P >0.05) between the inoculated and uninoculated seedlings under alkali stress(Fig. 4c).However,there were significant differences (P ≤0.05) between + ECMF and- ECMF treated seedlings when the concentration of NaHCO3was ≥10 mM (Fig. 4d). The results show that superoxide dismutase levels are more sensitive to alkali stress than to saline stress.
Content of osmotic adjustment substances
Most of the treatments affected the soluble protein contents of the seedlings (Fig. 5a, b; Table 4). There were no significant differences (P >0.05) between inoculated anduninoculated seedlings when Na2SO4was ≤10 mM and NaHCO3was 0 mM. There were significant differences(P ≤0.05) when the concentration of NaHCO3was ≥10 mM (Fig. 5b). In addition, the soluble protein content decreased when the concentration of NaHCO3was equal to 20 mM (Fig. 5b). In this research, the malondialdehyde(MDA) content increased significantly with increasing saline-alkali stress. However, the levels of MDA in the inoculated seedlings were lower than in the uninoculated(Fig. 5c, d). The results show that ectomycorrhizal inoculation can effectively reduce stress damage to P. sylvestrisvar. mongolica seedlings.
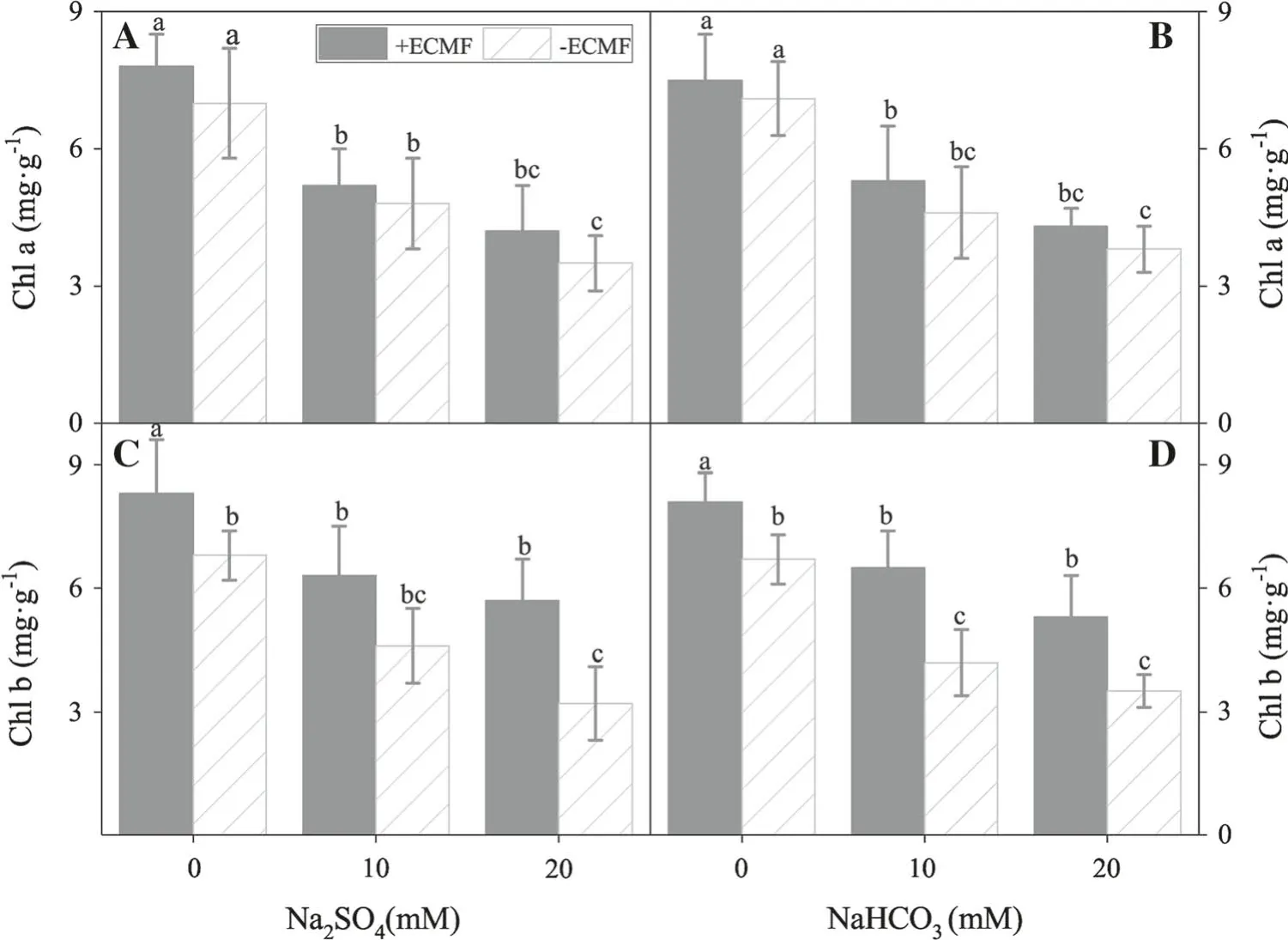
Fig. 3 Photosynthetic pigment content(mean ± SD,n = 10)as affected with or without ECMF under saline-alkali stress.Values with a different letter are significantly different for each treatment (P ≤0.05)

Table 2 Results of two-way ANOVA on chlorophyll a (Chl a) and chlorophyll b (Chl b)
Discussion
Soil salinization is a worldwide problem and can inhibit the germination of mycelia spores and inhibit their growth(Hirrel 1981; Pfeiffer and Bloss 1988; McMillen et al.1998). Becoming established is one of the most important and critical stages of seedling development under adverse Soilconditions such as salinity and alkalinity (Jones and Jones 1989; Yang et al. 2011). Salinity inhibits plant growth due to excess salt effects(Munns et al.2006).This study found a decrease in growth of P. sylvestrisvar.mongolica seedlings under high salinity, indicating that high saline soil inhibits mycelium growth and affects mycorrhizal colonization (Hajiboland et al. 2010). Inoculation with an ectomycorrhizal fungus was effective in reducing saline-alkali stress and its impact on the growth of P. sylvestrisvar. mongolica seedlings.
The symbiosis between an ectomycorrhizal fungus, in this instance S. luteus, and the plant can reduce the toxic effects of salt ions and effectively improve the tolerance of the plants to salt stress (Poss et al. 1985; Feng et al. 2002;Shen et al. 2004; Rabie 2005). Inoculation with ECMF significantly increased plant dry weight and became more obvious with increasing saline stress. This is consistent with the results obtained by Al-karaki and Rusan (2001).Under saline stress,ECMF improves plant dry weight,with above-ground biomass increasing more significantly than root biomass. This may be because, with ECMF colonization, carbohydrates are greater in above-ground parts than in roots (Latef and Chaoxing 2011).

Fig. 4 Antioxidant enzyme activity (mean ± SD, n = 10)as affected by inoculation with ECMF and under saline-alkali stress, values with different letters are significantly different for each treatment (P ≤0.05)

Table 3 Results of two-way ANOVA on CAT and SOD enzymes
In general, a saline-alkali environment decreases the chlorophyll content of seedlings. However, ECMF inoculation inhibited this reduction in P. sylvestrisvar. mongolica seedlings. In this study, chlorophyll a content in inoculated seedlings was not affected by saline stress when Na2SO4concentration was ≤10 mM; there were no significant differences (P >0.05) between the + ECMF and- ECMF treatments. This is consistent with results of previous studies (Wu et al. 2006). Thus, ectomycorrhizal fungi promote the growth of seedlings by improving their photosynthetic pigment levels and saline-alkali resistance.
Salt stress seriously affects the balance of free radical metabolism in plant cells, resulting in the accumulation of a large amount of reactive oxygen species (ROS), causing the peroxidation of cell membranes and destroying cell structure. Inoculation with an ectomycorrhizal fungus can regulate antioxidant enzyme activity and activate antioxidant enzyme scavenging free radicals.Under salinealkali stress, catalase (CAT) and superoxide dismutase(SOD) enzymes play important roles in defense against oxidative damage, regulate antioxidant enzymes mutual coordination between classes, eliminate intracellular ROS,and thus prevent cell membrane peroxidation.
In this study, saline-alkali stress affected antioxidant enzyme activities, and CAT and SOD activities in seedlings decreased with the degree of stress. Studies have found that the general stimulation of superoxide dismutase in plants is derived from mycorrhiza fungi(He et al.2007).Compared to plants without a mycorrhizal association,plants with mycorrhiza have higher antioxidant enzyme activities and lower lipid peroxidation accumulation, indicating that lower oxidative damage is associated with ectomycorrhizal association. Higher malondialdehyde(MDA) levels, as an index of cell membrane damage,indicate greater cell damage.
In this study,MDA levels of inoculated seedlings under saline-alkali stress are higher, indicating that cell membranes were damaged.MDA levels of inoculated seedlings under saline-alkali stress were always lower than those of the uninoculated seedlings, suggesting that inoculation is very effective at improving saline-alkali tolerance of seedlings and reducing MDA levels.
In conclusion, the ectomycorrhizal fungus, S. luteus,symbiosis increased the saline-alkali tolerance of P. sylvestrisvar. mongolica seedlings by increasing biomass,enhancing antioxidant enzyme activities, improving theadjustment of osmotic solutes, and enhancing chlorophyll contents.
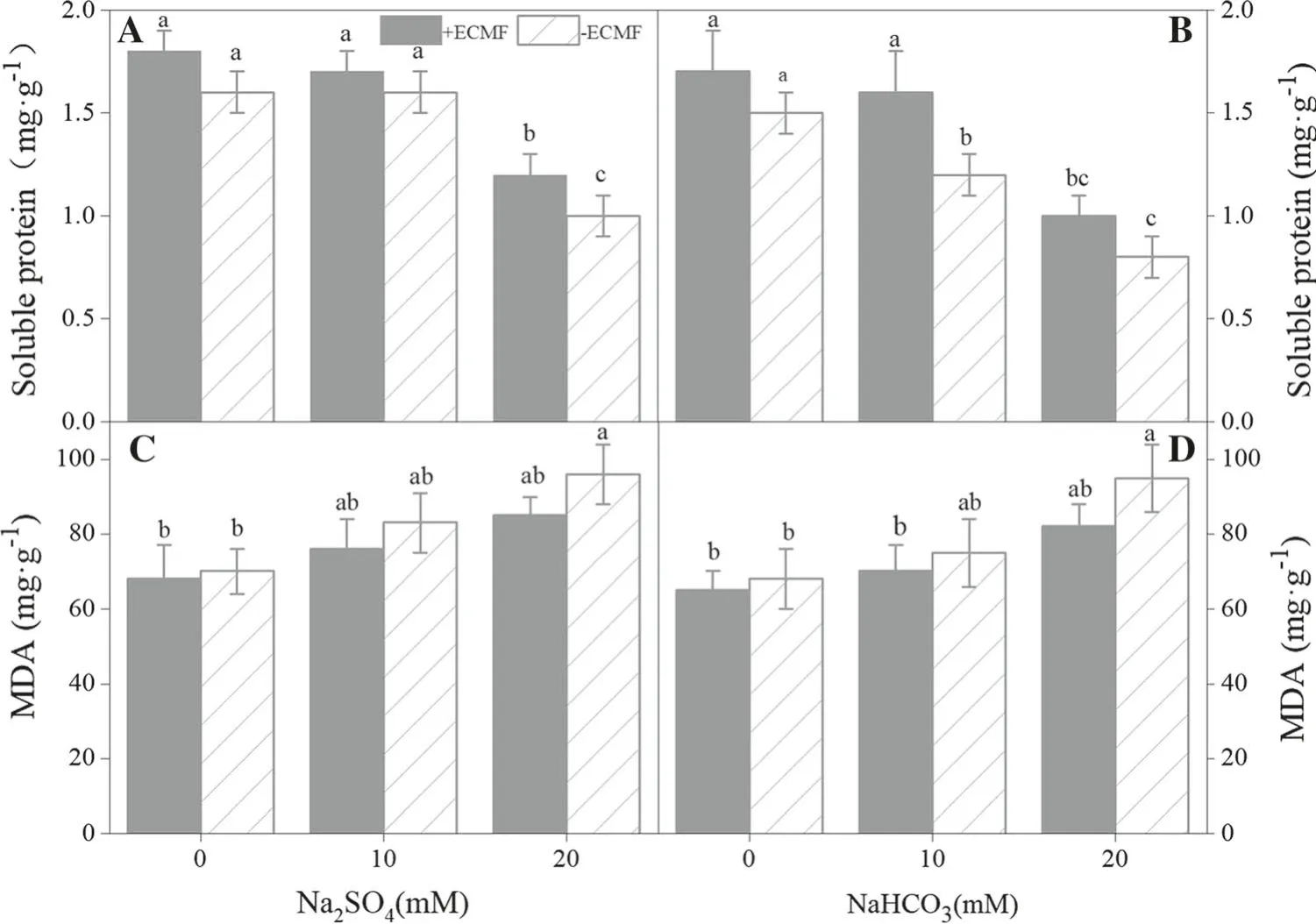
Fig. 5 Content of osmotic adjustment substances(mean + SD, n = 10) as affected by ECMF inoculation and under saline-alkali stress.Values with different letters are significantly different for each treatment (P ≤0.05)
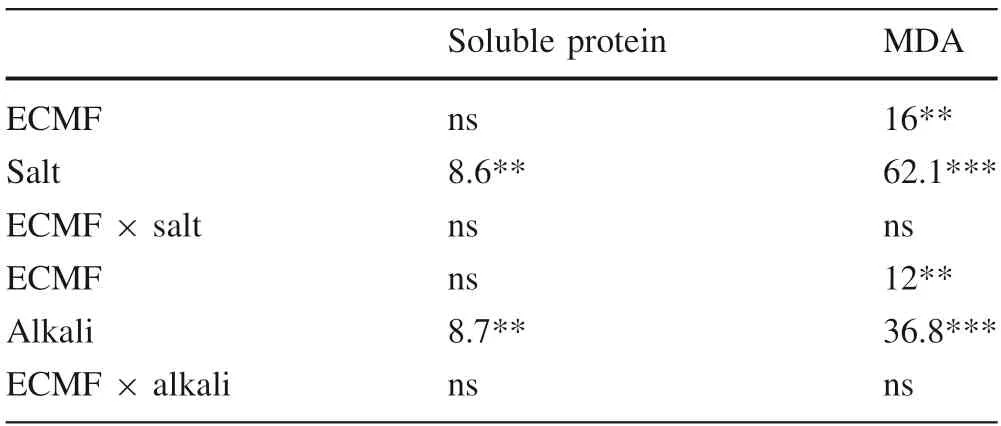
Table 4 Results of two-way ANOVA on soluble protein and MDA
In northeast China, soil salinization reduces tree survival. In this area, the use of inoculated P. sylvestrisvar.mongolica seedlings should be considered for the afforestation of degraded areas to help seedlings survive the saline-alkali environment.
杂志排行

Journal of Forestry Research的其它文章
- Do increasing respiratory costs explain the decline with age of forest growth rate?
- At what carbon price forest cutting should stop
- Mapping the risk of winter storm damage using GIS-based fuzzy logic
- Effects of seed moisture content, stratification and sowing date on the germination of Corylus avellana seeds
- Comparison of seed morphology of two ginkgo cultivars
- De novo assembly of the seed transcriptome and search for potential EST-SSR markers for an endangered, economically important tree species: Elaeagnus mollis Diels