雷州半岛红树林短吻鲾形态性状与体质量的相关性及通径分析
2020-05-13蔡润佳汤保贵张健东黄建盛潘传豪初庆柱王忠良
蔡润佳, 汤保贵, 陈 刚, 张健东, 黄建盛, 潘传豪, 初庆柱, 王忠良, 张 静
雷州半岛红树林短吻鲾形态性状与体质量的相关性及通径分析
蔡润佳, 汤保贵, 陈 刚, 张健东, 黄建盛, 潘传豪, 初庆柱, 王忠良, 张 静
(广东海洋大学 水产学院, 广东 湛江 524088)
为了研究雷州半岛红树林短吻鲾()各个形态特征与体质量的相关性, 对随机选取的153尾短吻鲾的全长(1)、体长(2)、头长(3) 、体高(4)、眼径(5)、眼间距(6)、吻长(7)、尾柄长(8)、尾柄高(9)、吻端至第一背鳍起点(10)、吻端至胸鳍起点(11)、吻端至腹鳍起点(12)、吻端至臀鳍起点(13)等13个指标和体质量()进行逐一测量, 根据所得数据整理出各个性状的表型参数, 计算各个指标与体质量以及不同指标两两之间的相关系数, 并分别求出以体质量()为因变量、其他性状(X)为自变量的通径系数以及决定系数。结果表明, 短吻鲾的13个形态性状对体质量的相关系数皆达极显著水平(<0.01); 在通径分析中, 短吻鲾仅体长(2)、吻长(7)、全长(1)3个形态性状对体质量数据的通径系数达到极显著水平(<0.01), 可将其确定为影响体质量的主要性状, 其中体长对体质量的直接影响最大(2=0.570); 通过逐步线性回归分析, 确定回归方程为:= –8.366+0.5702+0.1487+ 0.2661。决定系数显示, 体长对体质量的决定系数最大, 达到32.5%。各共同决定系数的总和∑为0.908, 大于0.85, 表明本研究中已经将影响短吻鲾体质量的主要形态性状全部纳入。此研究有助于确定并量化短吻鲾不同性状间的关系。
红树林; 短吻鲾(); 形态性状; 通径分析; 多元回归方程
红树林是热带和亚热带地区海湾以及河口泥滩上特有的以红树植物为主体的灌木和乔木群落, 环境较复杂, 营养来源广, 适合各种水生生物的生长、繁殖、栖息[1-4], 是世界上生物多样性最丰富、最具生产力的海洋生态系统之一[5]。由于其独特的生境, 红树林在减风抗浪[6]、保护堤坝[7]和维护生态平衡[8]等方面具有不可替代的作用。虽然红树林物种较丰富, 但由于靠近海岸易受人类活动的影响, 加之水质以及气温变化等原因, 90%以上的红树林均受到不同程度的影响。目前对红树林的保护主要集中在对其进行资源调查, 持续跟踪各个红树林区域的物种数量变化情况, 但未见对红树林的某个具体物种进行深入分析的报道。短吻鲾()隶属鲈形目、鲾科, 为我国南方常见小型暖水性鱼种, 主要栖息在近岸海区, 捕食小型甲壳类等为生。红树林为其生长繁殖提供了大量的食物和躲避、繁殖场所, 故在水温较高的红树林区域常有短吻鲾的分布。
通径分析是以相关分析及回归分析为基础的进一步研究。应用相关分析和通径分析的方法, 判定所研究物种各个外部形态性状与体质量之间的关联程度, 从而分别算出各个性状对体质量的直接和间接影响。此方法主要应用在对养殖生物的遗传选育上, 通过对外部形态性状的选择进而达到选种的目的, 有助于提高育种的工作效率[9-12]。将通径分析应用在红树林鱼类物种中, 可分析同个物种不同形态性状间的相关性, 对掌握红树林物种的表型相关数据有极大的补充。
本研究通过收集短吻鲾13个外部形态性状的数据, 利用相关和通径分析的方法对所收集的数据进行处理, 确定影响短吻鲾体质量的主要外部形态形状以及计算出各个形态形状对体质量的直接作用和间接作用的大小, 并且在偏回归系数的显著性检验中, 将检验不显著的形态性状剔除, 建立雷州半岛红树林短吻鲾形态性状与体质量间之间的最优多元线性回归方程, 为确定该物种影响体质量的主要形态性状及性状之间的相互关联提供依据。
1 材料与方法
1.1 实验材料
实验材料取自雷州半岛红树林, 一共四个季度, 五个地点(北潭、附城、高桥、和安、企水)采样所得并随机抽取其中的153尾短吻鲾, 将鱼体表水分擦干后, 参照孟庆闻等[13]鱼类的测量标准分别逐一测量其全长(1)、体长(2)、头长(3) 、体高(4)、眼径(5)、眼间距(6)、吻长(7)、尾柄长(8)、尾柄高(9)、吻端至第一背鳍起点(10)、吻端至胸鳍起点(11)、吻端至腹鳍起点(12)、吻端至臀鳍起点(13)等13个指标。用电子天平测量体质量。
1.2 数据分析
初步统计整理13个性状表型值的测定结果之后, 使用SPSS 19.0软件“Analyze-Descriptive Statitics- Explore”命令对所测量的全部体质量进行正态性检验, 对所采集的样本分别采用通径分析和回归分析[14]的方法, 再留取偏回归系数检验呈显著水平的自变量建立多元回归方程, 并进行拟合度检验。相关系数、决定系数以及通径系数的计算公式参考袁美云等[15]。多元线性回归方程模型为:

式中: b0为常数项, b1、b2、b3、…、bi为偏回归数, x1、x2、x3、…、xi为各偏回归系数相对应的自变量。利用SPSS 19.0软件进行显著性水平分析。数据对比结果默认设置为当P<0.05为差异显著, P<0.01为差异极显著。
注: 1. 全长; 2. 体长; 3. 头长; 4. 体高; 5. 眼径; 6. 尾柄长; 7. 尾柄高; 8. 吻端至胸鳍起点; 9. 吻长; 10. 眼间距; A. 吻端; C. 腹鳍起点; D. 臀鳍起点
2 结果与分析
2.1 短吻鲾全体样本的体长组成
统计结果显示, 所采集的153尾短吻鲾样本的全长范围为49~103 mm, 平均全长为71 mm, 体长范围为38~84 mm, 平均体长为56 mm。短吻鲾体长分布呈现6个范围(如图2所示), 其中体长范围50~ 60 mm为优势体长组, 占全体样品总数的47.7%。
2.2 生长相关性状表型参数
表1列出了短吻鲾13个形态性状和体质量表型统计量的结果。如表1所示, 在各形态性状表型参数测量结果中, 变异系数最大的是体质量(), 变异系数达到69.98%, 其次是吻端至腹鳍起点(12), 变异系数达60.92%, 变异系数最小的是眼径(5), 为19.07%,其他各形态性状指标的变异系数也均较大, 分别在19.15%~36.61%的区间内波动。

图2 短吻鲾全体样本的体长组成
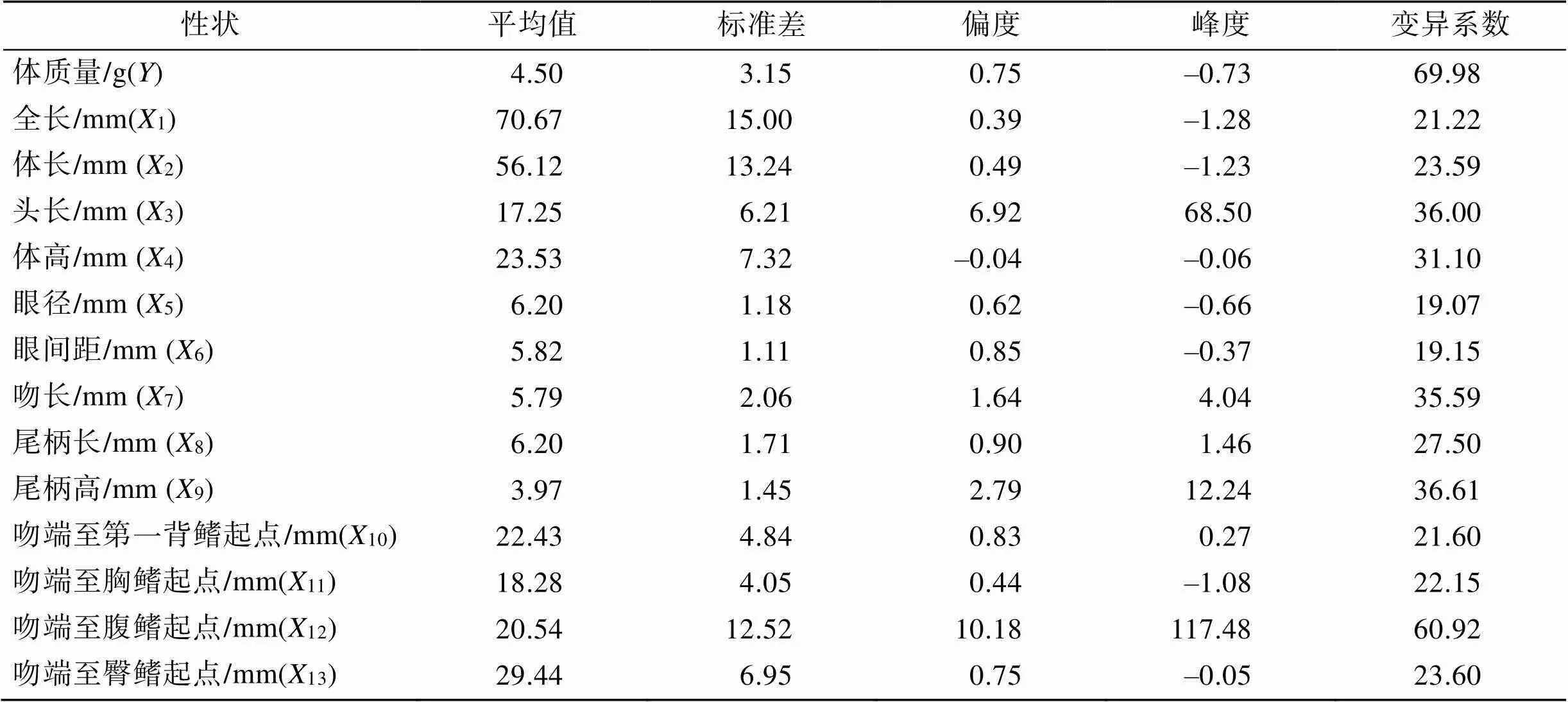
表1 短吻鲾各性状的表型统计量
2.3 性状间的相关系数
153尾短吻鲾各形态性状两两间以及各个性状与体质量的相关性数据显示(表2), 各形态性状间大部分呈极显著关系, 仅有12与3、8、10、11之间呈显著关系。其中1与2关系最大, 相关系数为0.971,3与12的关系最小, 相关系数为0.165。在各形态性状与体质量的相关系数中, 相关系数最大的是2, 达到0.947, 其次是1, 相关系数为0.935。12与的相关系数最小, 为0.262。
2.4 各形态性状对体质量影响的通径分析及作用分析
依据表型相关性分析和通径分析原理, 通过统计软件SPSS 19.0计算出各形态性状对体重的通径系数, 经显著性检验, 仅留全长、吻长、体长三个性状。由表3可知, 在这3个研究性状中, 对体质量的直接作用最为显著的是体长(2), 达到0.570, 其次是全长(1), 为0.266。而吻长(7)对体质量的直接作用最小, 仅为0.148。对体质量()的间接作用的影响大小依次为: 全长(1)>吻长(7)>体长(2)。由上述结果看出, 体长主要是通过直接作用来影响体质量的。
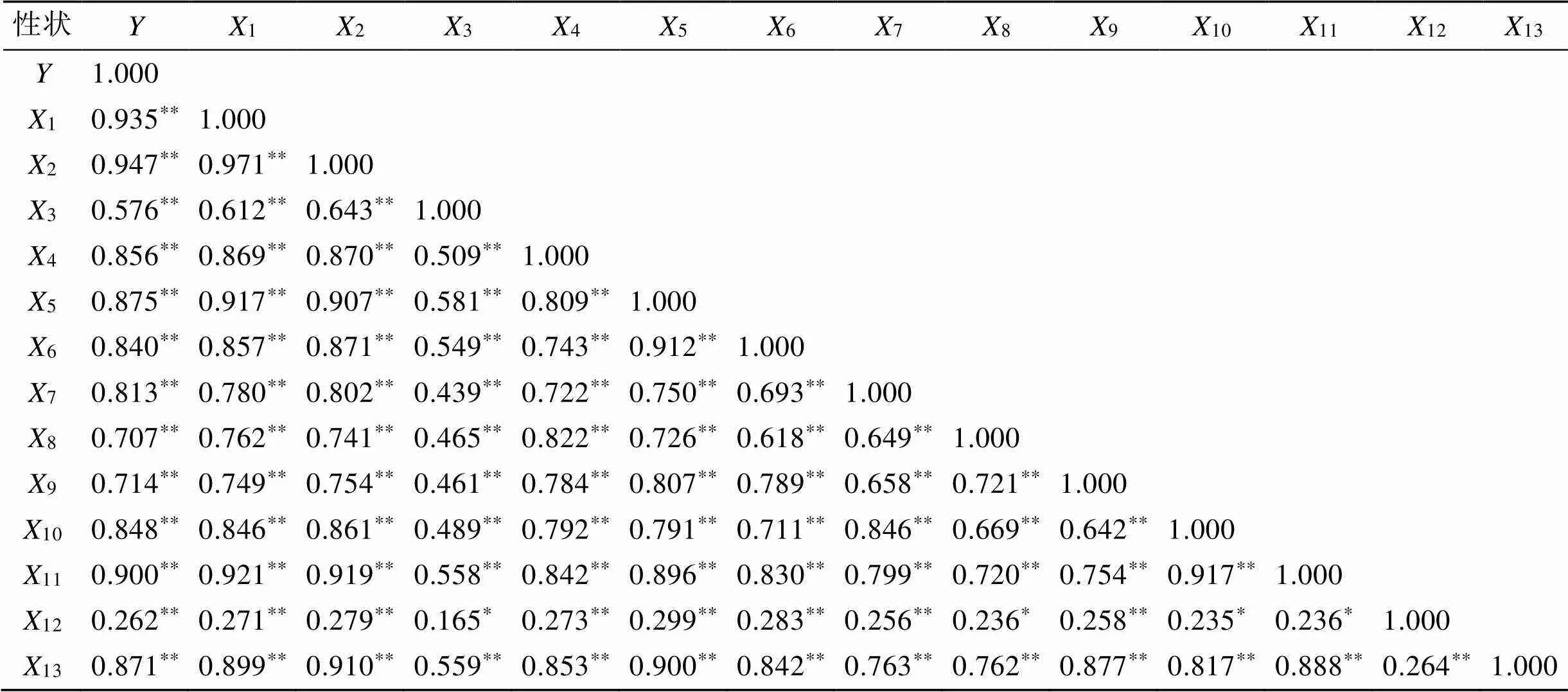
表2 短吻鲾各性状间的相关系数
注: **表示相关性极显著(<0.01), *表示相关性显著(<0.05)

表3 短吻鲾3个形态性状对体质量的影响
2.5 各形态性状对体质量的决定系数分析
在表4中, 位于表格对角线上的数值表示的是单个性状对体质量的决定系数, 而在对角线以上的部分, 表示的是两两性状对体质量的共同决定系数。体长(2), 吻长(7), 全长(1)对体质量的单独决定系数以及各共同决定系数相加, 得到总的决定系数∑为0.908(大于0.85), 表明影响短吻鲾体质量的主要形态性状已经全部确定。其中, 体长(2)对体质量的决定系数最大(0.325), 体长与全长的共同决定系数最大(0.294)。
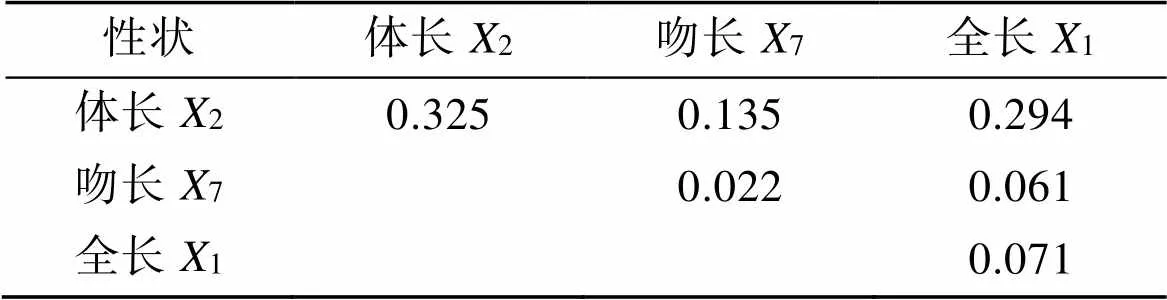
表4 短吻鲾3个形态性状对体质量的决定系数
2.6 各形态性状对体质量的复相关性
由表5可知, 随着将第3个自变量引入到方程中, 其复相关系数、相关指数和校正相关指数均达到最大, 其中复相关系数为0.935, 相关系数达到0.909, 校正相关指数达到0.907。标准误差达到最小, 仅为0.961。充分说明体长、吻长、全长是影响体质量的主要形态性状。表明所选的自变量已经囊括了对体质量绝大部分的影响。
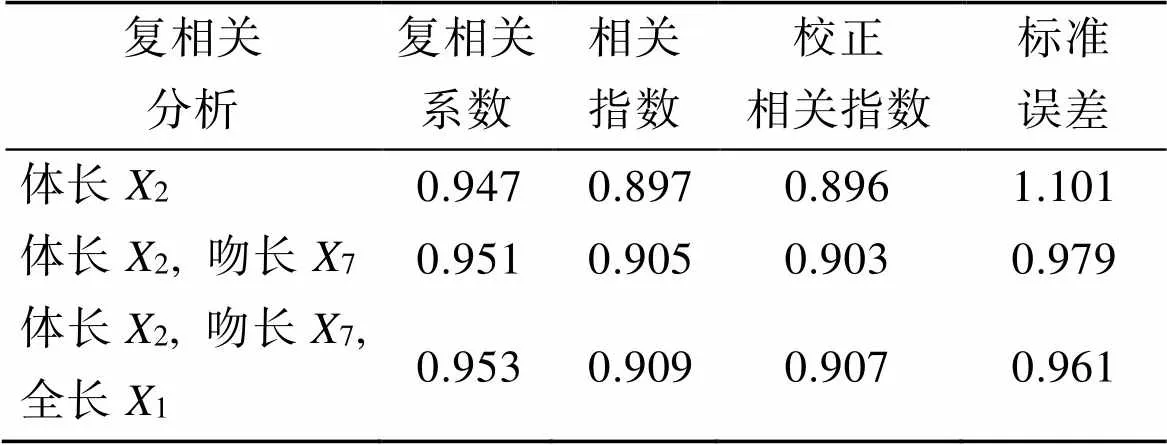
表5 短吻鲾3个形态性状对体质量的复相关系数
2.7 多元回归方程的建立及其回归分析
建立以测量样本活体体质量为因变量、以形态指标为自变量的回归方程如下:

式中,2、7、1分别为体长、吻长、全长。
如表6所示, 回归常数和所有的标准偏回归系数均达到极显著(<0.01)。方差分析结果(表7)表明, 回归关系达到极显著水平(<0.01)。经回归预测, 估计值与观察值不存在显著差异, 说明所列方程可以在实际运用过程中使用。
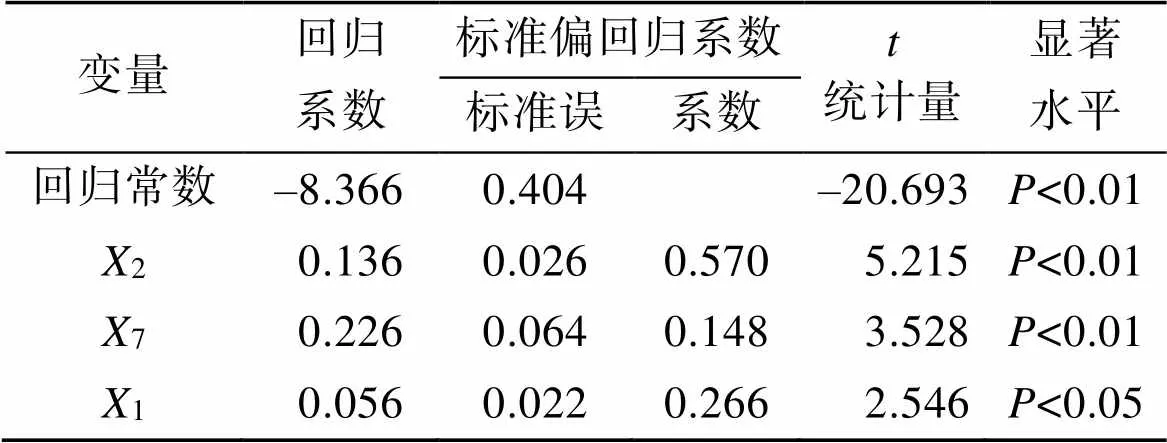
表6 偏回归系数和回归常数的显著性检验表

表7 多元回归方程的方差分析表
3 讨论
3.1 短吻鲾形态性状和体质量的相关性
对鱼类的进行通径分析中发现, 各个形态性状和体质量的相关性不尽相同。物种形态特征的差别是主要的因素之一。赵旺等[16]在5月龄的斜带石斑鱼()的研究中发现, 对体质量影响最大的4个形态性状分别是眼间距、尾柄高、体高和体宽; 杨月静等[17]研究表明, 体长、体高、体宽、吻长、尾柄长共5个形态性状均对13月龄的齐口裂腹鱼()体质量有较大的影响; 李玉全[18]指出, 半滑舌鳎()体质量主要与体高、体长、体厚和头长有较大关系。本研究对短吻鲾的分析中发现, 所测量的13个形态性状与体质量的相关性均达到了极显著水平(<0.01), 与体质量相关系数最大的3个性状分别是体长、吻长和全长。
通过对同个物种不同发育阶段的分析可得出, 影响体质量的主要形态性状可随着鱼体发育而发生改变。如翘嘴鳜()早期体质量主要受全长和体高两个性状的影响, 而随着鱼体的生长, 这两个性状对体质量的影响则逐渐减小[19]。1 龄洛氏鱥()影响体质量的4个性状分别是体长、体高、全长和尾柄高, 影响2龄鱼体质量的形态指标则是体长和体高[20]。耿绪云等[21]研究表明, 梭鱼()体长、体高是影响体重的共同性状, 在0、1、3、5龄组, 体长对体重的作用最大; 在2、4龄组, 体高对体重的作用最大。可见影响形态性状与体质量相关性的因素主要是物种形态间的区别和生长阶段的差异。
3.2 影响短吻鲾体质量的形态性状的确定
对鱼种进行表型性状和体质量的相关性分析, 虽能揭示各个性状分别与体质量的线性关系, 但不能准确的反映性状之间的相互关系, 也就不能准确全面地考察自变量与因变量之间的相关程度, 所以需要结合通径分析以及回归分析。在本研究中, 短吻鲾13个形态性状与体质量间的表型相关系数均达到了极显著水平(<0.01), 在各形态性状与体质量间的表型相关系数按降序的排列中, 以全长、吻端至胸鳍起点和体长三个变量位列前三, 但是在通径分析所得的检验结果却与表型相关分析结果出现差异。各形态性状中, 只有全长、吻长和体长这三个变量对体质量性状的通径系数达到极显著水平(<0.01)。因此回归方程仅纳入全长、吻长和体长3个性状, 剔除其他10个表型性状。比较3个性状对体质量的直接或间接作用可知, 全长对体质量的直接作用大于间接作用, 而吻长和体长两个性状的间接作用远大于直接作用。以这3个性状为自变量, 体质量为因变量建立回归方程, 结果发现, 体长、吻长和全长三个形态性状对体质量的总决定系数为0.908, 超过0.85[22], 表明此回归方程已经将对短吻鲾体质量产生影响的主要形态性状全部列出, 并可确定影响短吻鲾体质量的主要形态性状为全长、吻长和体长。全长是奥里亚罗非鱼()4月龄雄鱼[23], 1月龄和4月龄的卵形鲳鲹()[24]等鱼种影响体质量的主要性状之一, 体长是尼罗罗非鱼()[25], 半滑舌鳎[18]等鱼种的主要因素之一。在目前的研究中, 未见吻长对体质量产生主要影响的鱼种, 这可能跟短吻鲾的外部形态特征有关。
[1] Odum E P, Heald R J. Detritus Based Food Wed of an Estuarine Mangrove Community[M]. New York: Academic Press, 1975: 256-286.
[2] Thayer W G, Colby D R, Hetller W F. Utilization of the red mangrove prop root habitat by fishes in south Florida[J]. Marine Ecology Progress Series, 1987, 35: 25-38.
[3] Vance D J, Haywood M D E, Heales D S, et al. How far do prawns and fish move into mangrove? Distribution of juvenile banana prawnsand fish in a tropical mangrove forest in northern Australia[J]. Marine Ecology Progress Series, 1996, 131: 115-124.
[4] Robertson A, Duke N C. Mangroves as nursery sites: comparisons of the abundance and species composition of fish and crustaceans in mangroves and other near shore habitats in tropical Australia[J]. Marine biology, 1987, 96: 193-205.
[5] 林鹏.中国红树林生态系统[M]. 北京: 科学出版社, 1997: 1-10. Lin Peng. Chinese mangrove ecosystem[M]. Beijing: Science Press, 1997: 1-10.
[6] Kathiresan K, Rajendran N. Coastal mangroveforests mitigated tsunami[J]. Estuarine, Coastaland Shelf Science, 2005, 65(3): 601-606.
[7] 林鹏. 中国红树林湿地与生态工程的几个问题[J]. 中国工程科学, 2003, 5(6): 33-38. Lin Peng. The characteristics of mangrove wetlands and some ecological engineering questions in China[J]. Engineering Science, 2003, 5(6): 33-38.
[8] 林益明, 林鹏. 福建红树林资源的现状与保护[J]. 生态经济, 1999, 3: 16-19. Lin Yiming, Lin peng. The status and conservation of the mangrove resources in Fujian[J]. Ecological Economy, 1999, 3: 16-19.
[9] 董世瑞, 孔杰, 万初坤, 等. 中国对虾形态性状对体重影响的通径分析[J]. 海洋水产研究, 2007, (3): 15-22.Dong Shirui, Kong Jie, Wan Chukun, et al. Path analysis of effects of morphometric attributes on body weight of[J]. Marine Fisheries Research, 2007, (3): 15-22.
[10] 黄建盛, 张敬威, 陈刚, 等. 褐点石斑鱼(♀)×清水石斑鱼(♂)杂交子代幼鱼形态性状对体质量影响的通径分析[J]. 广东海洋大学学报, 2017, 37(3): 23-28. Huang Jiansheng, Zhang Jingwei, Chen Gang, et al. Path analysis of morphometric traits on body weight for hybrid grouper (♀×♂)[J]. Journal of Guangdong Ocean University, 2017, 37(3): 23-28.
[11] 刘贤德, 蔡明夷, 王志勇, 等. 不同生长时期大黄鱼形态性状与体重的相关性分析[J]. 热带海洋学报, 2011, 29(5): 159-163. Liu Xiande, Cai Mingyi, Wang Zhiyong, et al. Correlation analysis of morphometric traits and body weight of large yellow croakerat different growth stage[J]. Journal Of Tropical Oceanography, 2011, 29(5): 159-163.
[12] 黄建盛, 陈刚, 张健东, 等. 褐点石斑鱼不同月龄形态性状的主成分及通径分析[J]. 水产学报, 2017, 41(7): 1105-1115. Huang Jiansheng, Chen Gang, Zhang Jiandong, et al. Principal component and path analysis of morphological traits ofat different month ages[J]. Journal of Fisheries of China, 2017, 41(7): 1105- 1115.
[13] 孟庆闻, 苏锦祥, 缪学祖. 鱼类分类学[M]. 北京: 中国农业出版社, 1995: 29-30. Meng Qingwen, Su Jinxiang, Miao Xuezu. Fish Taxonomy[M]. Beijing: China Agriculture Press, 1995: 29- 30.
[14] 杜家菊, 陈志伟. 使用SPSS线性回归实现通径分析的方法[J]. 生物学通报, 2010, 45(2): 4-6. Du Jiaju, Chen Zhiwei. Method for realizing path analysis using SPSS linear regression[J].China Academic Journal Electronic Publishing House, 2010, 45(2): 4-6
[15] 袁美云, 刘双凤, 韩志忠. 3月龄施氏鲟形态性状对体质量的影响分析[J]. 中国水产科学, 2010, 17(3): 507-513. Yuan Meiyun, Liu Shuangfeng, Han Zhizhong. Mathematical analysis of morphometric attribute effects on body weight for three-month-old[J]. Journal of Fishery Sciences of China, 2010, 17(3): 507-513.
[16] 赵旺, 杨蕊, 胡静, 等. 5月龄斜带石斑鱼形态性状对体质量影响的通径分析[J]. 大连海洋大学学报, 2017, 32(5): 557-562. Zhao Wang, Yang Rui, Hu Jing, et al. Path analysis of morphological traits effects on body weightof five- month-old grouper[J]. Journal of Dalian Ocean University, 2017, 32(5): 557-562.
[17] 杨月静, 向梦斌, 刘庭, 等. 13月龄养殖齐口裂腹鱼形态性状与体质量的关系[J]. 河南农业科学, 2019, 48(6): 139-144. Yang Yuejing, Xiang Mengbin, Liu Ting, et al. Relationship between morphological traits and body mass of cultured 13-month-old[J]. Journal of Henan Agricultural Sciences, 2019, 48(6): 139-144.
[18] 李玉全. 线性回归法实现半滑舌鳎幼鱼表型性状与体重的通径分析[J]. 生物数学学报, 2014, 29(1): 185- 191. Li Yuquan. Path Analysis between morphometric attributes and body weight on juvenileby multiple regression[J]. Jouenal of Biomathematics, 2014, 29(1): 185-191.
[19] 张进, 梁旭方, 郭小泽, 等. 翘嘴鳜形态性状对体质量的影响[J]. 华中农业大学学报, 2013, 32(5): 100-105. Zhang Jin, Liang Xufang, Guo Xiaoze, et al. Mathematical analysis of effects of morphometric traitson body weight of[J]. Journal of Huazhong Agricultural University , 2013, 32(5): 100-105.
[20] 张永泉, 白庆利, 郭文学, 等. 洛氏鱥形态性状对体质量的影响[J]. 生态学杂志, 2013, 32(11): 3063-3068. Zhang Yongquan, Bai Qingli, Guo Wenxue, et al. Effects of morphological traits on body mass of[J]. Chinese Journal of Ecology 2013, 2013, 32(11): 3063-3068.
[21] 耿绪云, 马维林, 李相普, 等. 梭鱼()外部形态性状对体重影响效果分析[J]. 海洋与湖沼, 2011, 42(4): 530-537. Geng Xuyun, Ma Weilin, Li Xiangpu, et al. Morphometric attributes to body weight for the redlip mullet[J]. Oceanologia et Limnologia Sinica, 2011, 42(4): 530-537.
[22] 齐明, 侯俊利, 楼宝, 等. 一龄四指马鲅形态性状对体重的影响分析[J]. 浙江海洋学院学报(自然科学版), 2014, 33(2): 134-139. Qi Ming, Hou Junli, Lou Bao, et al. Effects of Morphometric Attributes on Body Weight for One-year-old[J]. Journal of Zhejiang Ocean University (Natural Science), 2014, 33(2): 134- 139.
[23] 曾兰, 林勇, 张永德, 等.奥利亚罗非鱼形态性状与体重的通径分析[J]. 西南农业学报, 2012, 25(1): 295- 301. Zeng Lan, Lin Yong, Zhang Yongde, et al. Path analysis on mcorphological rraits and body weight of[J]. Southwest China Journal of Agricultural Sciences, 2012, 25(1): 295-301.
[24] 区又君, 吉磊, 李加儿, 等. 卵形鲳鲹不同月龄选育群体主要形态性状与体质量的相关性分析[J]. 水产学报, 2013, 37(7): 961-969. Ou Youjun, Ji Lei, Li Jiaer, et al. Correlation analysis of major morphometric traits and body weight ofselective group at different month ages of[J]. Journal of Fisheries of China, 2013, 37(7): 961-969.
[25] 唐瞻杨, 肖俊, 李莉萍, 等. 尼罗罗非鱼()不同月龄性状的主成分与判别分析[J]. 海洋与湖沼, 2012, 43(2): 288-293. Tang Zhanyang, Xiao Jun, Li Liping, et al. Principal component and discriminant analyses of traits of nile tilapia () at different ages[J]. Oceanologia et Limnologia Sinica, 2012, 43(2): 288-293.
Correlation and path coefficient analysis of the body weight and morphometric traits ofin Leizhou Peninsula
CAI Run-jia, TANG Bao-gui, CHEN Gang, ZHANG Jian-dong, HUANG Jian-sheng, PAN Chuan-hao, CHU Qing-zhu, WANG Zhong-liang, ZHANG Jing
(Fisheries College, Guangdong Ocean University, Zhanjiang 524088, China)
The correlations between various morphological characteristics and the body weight ofof the Leizhou Peninsula in South China were investigated by measuring the full length (1), body length (2), head length (3), body height (4), eye diameter (5), eye spacing (6), kiss length (7), tail handle length (8), tail handle height (9), kiss end to first dorsal fin starting point (10), kiss end to pectoral fin starting point (11), kiss end to beginning of the pelvic fin (12), start of kiss to anal fin (13), and body weight () of 153 randomly selected individuals and sorting the phenotypic parameters of each trait according to the data obtained. The correlation coefficients between each of the 13 morphological traits and body weight and among the various morphological traits were then calculated, and the path coefficient and coefficient of determination were determined using the body weight () as the dependent variable and the other traits (X) as the independent variables. It was found that the correlations between each morphological trait and body weight were extremely significant (< 0.01). However, in the path analysis, only the full length (1), body length (2), and kiss length (7) were highly significantly related to body weight (< 0.01), indicating that these are the main traits that affect body weight, with stepwise linear regression analysis giving a regression equation of= −8.366 + 0.5702+ 0.1487+ 0.2661. Among these, body length had the largest direct effect (2= 0.570) and also had the largest coefficient of determination for body weight, reaching 32.5%. The sum of the common decision coefficients ∑was 0.908, which is greater than 0.85, indicating that all of the major morphological traits that affect the weight ofwere included in this study. These findings help to determine and quantify the relationship between the various traits ofand to understand the degree of association between them.
mangrove forest;; morphological trait; path analysis; multiple regression equation
Oct. 23, 2019
[Survey of benthic resources in Zhanjiang Mangrove National Nature Reserve, Guangdong Province, No. B16377; Investigation of benthic resources of fish and crustaceans in Zhanjiang Mangrove National Nature Reserve, No. B16378]
S965
A
1000-3096(2020)04-0052-07
10.11759/hykx20191023001
2019-10-23;
2020-01-22
广东湛江红树林国家级自然保护区底栖生物资源调查(B16377); 湛江红树林国家级自然保护区鱼类与甲壳类资源调查(B16378)
蔡润佳(1995-), 男, 广东汕头人, 硕士研究生, 研究方向为水产经济动物生物学, E-mail: 906636294@qq.com; 汤保贵(1975-),
, 博士, 副教授, 主要研究方向为鱼类养殖与遗传育种, E-mail: zjtbg@163.com
(本文编辑: 康亦兼)
