褐飞虱ABCB基因的克隆与表达分析
2020-05-11闸雯俊李三和陈志军杨国才徐华山李培德游艾青
闸雯俊,李三和,周 雷,陈志军,刘 凯,杨国才,徐华山,李培德,游艾青,2*
(1.湖北省农业科学院粮食作物研究所,粮食作物种质创新与遗传改良湖北省重点实验室,武汉 430064;2.长江大学,主要粮食作物产业化湖北省协同创新中心,荆州 434025)
所有的细胞和细胞器都是通过脂质膜与外界环境分离的,需要转运蛋白来转运这些膜上的多种化合物(Higgins CF,1992)。以下事实证明了膜转运的重要性:在大肠杆菌和人类中,约10%和4%的基因编码转运相关的蛋白(Blattneretal.,1997)。大多数ABC蛋白作为主要的活性转运蛋白发挥作用,需要结合和水解ATP来转运脂质膜上的底物,其转运的底物包括糖、氨基酸、金属离子、多肽、蛋白质、细胞代谢产物和药物等(王华丙等,2007)。ABC转运蛋白最早发现于细菌,是细菌质膜上的一种具有运输功能的ATP酶,由两个结合和水解ATP的胞质核苷酸结合域(NBD)和两个完整的跨膜域(TMD)组成。NBD域包含几个保守序列,如Walker A (P-loop)、Walker B和特征基序C(吴转斌和吴金美,2008)。跨膜结构域由5~6个跨膜螺旋组成,并提供底物特异性。功能性转运体的4个结构域可以存在于一种蛋白质中,也可以分布在多种蛋白质上。在后者的情况下,ABC蛋白需要同二聚体或异二聚体来形成功能性ABC转运体(Higgins and Linton,2004)。根据NBD的序列相似性,ABC蛋白家族被分为8个亚家族,用字母a~h表示。
褐飞虱Nilaparvatalugens属同翅目飞虱科昆虫,又称褐稻虱。褐飞虱是水稻主要害虫之一,在中国长江流域和华南及西南广大稻区发生频繁、危害严重(王传之等,2008)。本研究从褐飞虱体内分离得到ABCB基因,分析了ABCB基因的全长、结构,为研究褐飞虱ABCB基因的功能奠定了基础。
RNA干扰是指在进化过程中高度保守的、由双链RNA(double-stranded RNA,dsRNA)诱发的、同源mRNA高效特异性降解的现象。高浓度的dsRNA才能保证靶标基因的连续抑制,利用体外合成的dsRNA来喂养昆虫成本相对昂贵。HT115(DE3)菌株是一类特殊的RNase III缺陷型大肠杆菌菌株,可以饲喂线虫,主要用于秀丽隐杆线虫Caenorhabditiselegans的RNAi干扰试验。该菌株可以在LB或2YT培养基中正常生长,Tn10转座子的存在使其具有四环素抗性。该菌株染色体整合了λ噬菌体DE3区(DE3区含有T7噬菌体RNA聚合酶,在IPTG存在时可诱导T7 RNA聚合酶大量表达,进而启动线虫dsRNA的表达),可同时表达T7 RNA聚合酶和大肠杆菌RNA聚合酶,除了用于线虫RNAi干扰试验,也可用于pET系列、pGEX、pMAL等质粒的蛋白表达。利用大肠杆菌表达dsRNA已在粘虫Mythimnaseparate(Ganbaataretal.,2017)中得以实现。
利用dsRNA喂养是目前最方便的方法之一,这种方法已经在从属于7个目15种的昆虫物种中实现了。通过喂养,对于肠道的靶标基因表现出极高的沉默效率。对于其他组织的靶标基因而言,RNAi的效率会因为靶标基因的不同和昆虫种属的不同而差别很大。例如蜜蜂Apismellifera脂肪体的基因,通过喂养会诱导90%的卵黄蛋白原表达下降(Nunes and Simoes,2009);以及美洲散白蚁Reticulitermesflavipes调控Hexamerin储存蛋白表达下降50%~70% (Zhouetal.,2008);但是对于刺舌蝇Glossinamorsitans的铁转运蛋白没有任何的效果(Walsheetal.,2009)。同时喂养效率也受昆虫生长发育的影响。例如利用nitropin 2 dsRNA处理长红锥蝽Rhodniusprolixus的4龄若虫没有效果,但利用2龄若虫产生了42%的沉默效果(Araujoetal.,2006)。虽然利用dsRNA喂养的方法是非侵入性和方便的方法,不过喂养时dsRNA的摄取量和RNAi的效率会因为个体之间的差异而变得困难(Turneretal.,2006)。
因此,利用此技术通过人工喂养dsRNA沉默靶标而达到控制褐飞虱的目的。本研究通过构建NlABCB-dsRNA载体,转化E.coliHT115后,经IPTG诱导,形成NlABCB-dsRNA,喂养褐飞虱后,褐飞虱靶标基因抑制,存活率下降。
1 材料与方法
1.1 材料
1.1.1材料
褐飞虱品种为武汉大学饲养,饲养在感虫水稻品种台中1号TN1植株上,环境条件为25±2℃,相对湿度为80%,光照周期为L ∶D=16 ∶8。
1.1.2试剂
试验所用大肠杆菌E.coliDH5α,ExTaq酶,逆转录试剂盒和pMD18-T克隆载体,5′-Full RACE和3′-Full RACE试剂盒购自Takara公司。引物合成及序列测定由北京擎科新业生物技术有限公司完成。
1.2 方法
1.2.1总RNA的提取
采用TRIzol法抽提褐飞虱样品的总RNA,具体操作参考Invitrogen公司TRIzol® reagent试剂说明书。
1.2.2cDNA克隆
利用赤拟谷盗Triboliumcastaneum的ABC transporter蛋白(GenBank登录号:XP_971735.1)褐飞虱的ESTs数据库(http://bphest.dna.affrc.go.jp/)中进行TBLASTN查询,得到相应的相应EST片段。设计RACE外围引物3′RACE outer和5′RACE outer以及内围引物3′RACE inner和5′RACE inner。3′RACE和5′RACE模板的合成具体按照BDSMART RACE cDNA Amplification Kit User Manual说明书步骤操作。利用巢式PCR技术分别扩增褐飞虱基因3′端和5′端。或查询褐飞虱基因组,结合Augustus、FgeneSH等基因预测软件分析,设计引物直接从两端扩增全长cDNA。
1.2.3实时荧光定量PCR检测NlABCB转录差异
取-80℃保存不同生长发育时期褐飞虱和褐飞虱不同组织总RNA定量,按照反转录试剂盒说明书进行反转录获得褐飞虱的cDNA。在Roteogene 6000TM定量PCR仪(Corbett research)上进行10 μL体系的PCR反应,反应条件如下:94℃预变性2 min,然后以30~40个循环进行PCR扩增(94℃ 20 s,55℃ 15 s,72℃ 20 s)每个样品3个重复,取平均值。以褐飞虱Actin作为参照基因,采用2-ΔΔCt方法(Livak and Schmittgen,2001),比较不同样品基因表达水平。
1.2.4生物信息学分析
测序拼接获得cDNA全长后,利用ExPASy Translate tool软件(http://web.expasy.org/translate/)对翻译得到相应cDNA编码框进行预测,再利用下列在线工具进行蛋白质序列的功能域及结构域的预测:同源序列比对利用Clustalw(http://www.genome.jp/tools/clustalw/);进化树图的绘制利用MEGA 7.0软件;分子量和等电点分析利用ExPASy Compute pI/Mw tool (http://web.expasy.org/compute_pi/);二级结构的预测利用(http://npsa-pbil.ibcp.fr/)。
1.2.5人工喂养dsRNA
取2龄褐飞虱若虫用人工饲料作适应性预饲养,待若虫发育至目标基因高表达阶段,向人工饲料中添加dsRNA,每头幼虫喂养10 μg(1 μg/μL)dsRNA。RNA干扰设置2~3个浓度,同时设置空白对照组(纯人工饲料)和dsGFP对照组(含dsGFP的人工饲料)。处理时,每饲喂装置接入20头发育一致的褐飞虱,设置3个重复。考察指标包括靶标基因表达量、个体存活率、生长发育状况等。
2 结果与分析
2.1 褐飞虱NlABCB基因的克隆
以该EST序列为模板设计RACE引物,克隆得到NlABCB基因的5′cDNA片段和3′cDNA片段。5′cDNA序列长度约为0.9 Kb,3′cDNA序列长度约为0.8 Kb。全长序列经过测序拼接后,全长为1 468 bp,含有一个编码153个氨基酸的开放开放阅读框(图1)。
2.2 NlABCB蛋白分析鉴定
NlABCB 基因编码153个氨基酸。应用Computer pI/Mw Tool软件(http://web.expasy.org/compute_pi/)分析预测 NlABCB蛋白的分子量为16.8 kDa,等电点为8.8。二级结构预测分析发现,NlABCB 蛋白α螺旋占44.08%,延伸链占21.05%,β折叠占9.87%,随机卷曲占25%(图2)。
通过NCBI的Blastp(http://blast.ncbi.nlm.nih.gov/Blast.cgi)程序进行在线蛋白质序列比对,结果表明该基因与其它昆虫的ABCB蛋白序列相似性大于72.54%(表2)。例如与灰飞虱Laodelphaxstriatella的同源性为93.42%,最高;与美洲鲎Limuluspolyphemus的同源性为72.54%,最低。由此可以看出褐飞虱 NlABCB 与其它昆虫的ABC转运蛋白序列相似性较高,其序列相当保守。

表1 引物序列Table 1 Primer sequence

图1 NlABCB 的核苷酸和推测氨基酸序列Fig.1 Nucleotide and deduced amino acid sequence of NlABCB

表2 褐飞虱NlABCB基因与其它昆虫NlABCB基因氨基酸相似性比较Table 2 Amino acid similarities of NlABCB gene of Nilaparvata lugens and other insects
2.3 褐飞虱 NlABCB 基因不同生长发育时期和组织的分析
为了解NlABCB基因在褐飞虱不同生长发育阶段虫体中和虫体不同组织中的转录情况,分别提取1龄至5龄发育至24 h的若虫和雌雄成虫虫体的总RNA以及5龄若虫的中肠、唾液腺、脂肪体、表皮、腿和头部组织的总RNA,利用实时荧光定量PCR法检测该基因在褐飞虱不同发育阶段虫体中和不同组织的表达情况。试验结果表明,该基因在雄成虫的表达量较高(图3)。在不同组织中,在中肠的表达量最高(图4)。

图3 NlABCB 基因在不同生长发育时期的表达Fig.3 Expression of NlABCB at different growth and development stages
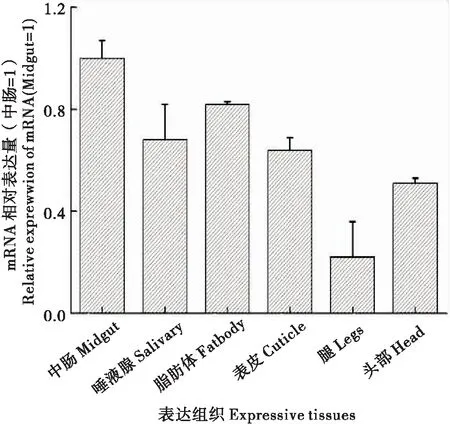
图4 NlABCB 基因在不同组织的表达分布Fig.4 Expression and distribution of NlABCB in different tissues
2.4 褐飞虱L4440-NlABCB-dsRNA的表达及其RNAi效果
以褐飞虱cDNA为模板,扩增褐飞風ABCB基因片段。Nco I和Sma I双酶切L4440载体后,转入菌株HT115后,IPTG便可表达dsRNA(图5)。分别让褐飞虱取食NlABCB-dsRNA进行RNAi。首先检测褐飞虱取食dsRNA后,对其体内NlABCBmRNA表达水平的影响。结果发现:褐飞虱取食dsRNA后,第2天和第4天分别抑制了25%和48%。因此,人工喂养dsRNA可以抑制靶基因表达,效果显著(图6)。同时,褐飞虱取食dsRNA后,致死效果明显。取食NlABCB-dsRNA后,其存活率显著低于对照组,对照组和试验组1 d、2 d、3 d、4 d的存活率分别为88.77%和86.67%、76.9%和70.78%、65.75%和55.67%、56.72%和46.69%。因此NlABCB基因可以进一步应用于褐飞虱抗虫(图7)。

图5 褐飞虱 NlABCB 的dsRNA电泳图Fig.5 The dsRNAs produced by the BPH NlABCB gene

图6 褐飞虱取食dsRNA后 NlABCB mRNA的表达Fig.6 Expression of NlABCB mRNA in Nilaparvata lugens after feeding on dsRNA
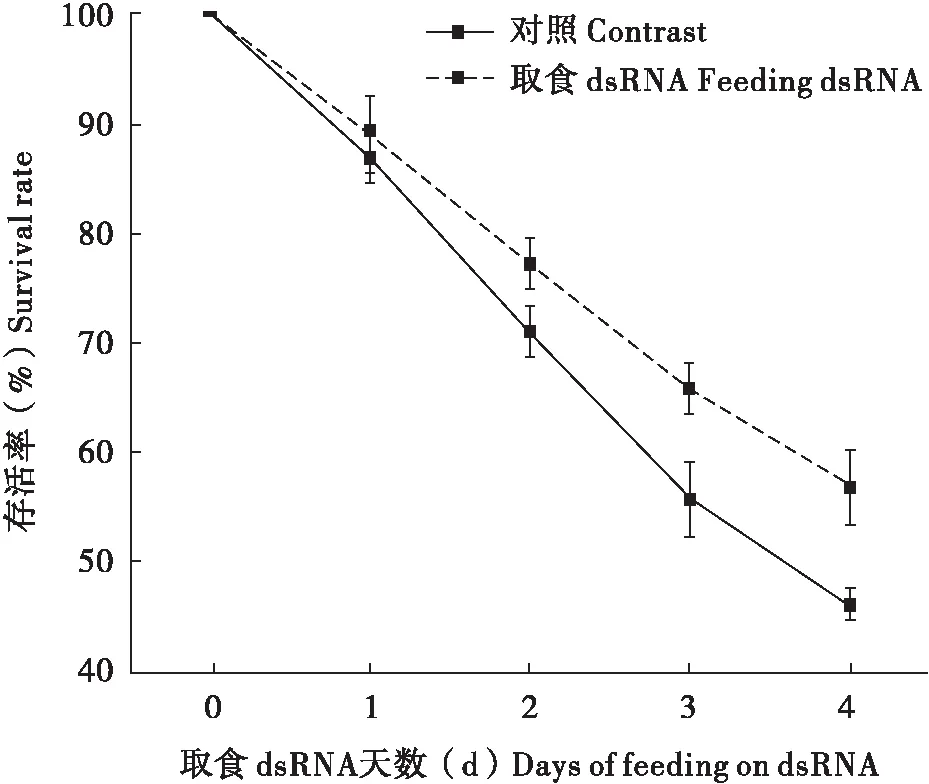
图7 褐飞虱取食dsRNA后存活率Fig.7 Survival rate of brown planthopper after feeding on dsRNA
3 结论和讨论
ABCB亚家族由半个(HTS)和整个转运体(FTS)组成。黑腹果蝇Drosophilamelanogaster中的ABCB-FTS已经被广泛研究,MDR65(ABCB)通过创建血红素屏障来对果蝇大脑进行化学保护(Mayeretal.,2009),MDR49(ABCB)在生殖细胞迁移中起着至关重要的作用(Ricardo and Lehmann,2009)。此外,研究表明果蝇接触到多环芳烃会诱导体内的ABCB FT/P-gfp表达(Vachetal.,2006)。同时给果蝇食物中添加含有0.1 mmol的抗叶酸药物甲氨蝶呤后,在果蝇成虫的马氏管和肠道中,MDR49(ABCB)、MDR50(ABCB)的表达会显著上调。另一方面,Broehan等人发现给蛹前期的赤拟谷盗注射ABCB- dsRNA后,赤拟谷盗的蛹成虫期会有严重的发育缺陷,从而致死;同时卵巢不能产卵导致雌性不育(Broehanetal.,2013)。
最近的一些成功的例子表明dsRNA喂养可用于作为一种潜在的防治害虫方式。Baum等通过筛选玉米根虫DiabroticavirgiferavirgiferaLeConte 的cDNA文库鉴定了290个编码重要蛋白的靶标基因,发现52 ng/cm2浓度的dsRNA可以产生明显的死亡率和发育迟缓(Baumetal.,2007)。通过取食获得RNAi的效果已在多种昆虫中得以实现,如苹果褐卷蛾Pandemisheparana(Turneretal.,2006),美洲散白蚁(Zhouetal.,2008),长红锥蝽(Araujoetal.,2006)等当中都有发现,所以这已经成为一种控制害虫的新方法。
本研究克隆得到NlABCB基因的全长,并且将氨基酸序列与其他昆虫的ABCB进行序列比对,发现与灰飞虱、湿木白蚁的ABCB蛋白在进化上亲缘关系较近。并且对褐飞虱ABCB蛋白序列分析发现,该蛋白α螺旋占44.08%,延伸链占21.05%,β折叠占9.87%,随机卷曲占25%。同时鉴定了NlABCB基因在褐飞虱不同生长发育阶段虫体中和虫体不同组织中的转录情况,该基因在雌性成虫的表达量较高。在不同组织中,在中肠的表达量最高。中肠组织是昆虫肠道组织中唯一与细胞表面直接接触的。同时有研究表明家蚕Bombyxmori中的ABCB基因是特异表达的,在蜕皮和化蛹期间受20E正调节(Liuetal.,2011)。同时构建L4440-NlABCB的dsRNA载体,转化E.coliHT115后,经IPTG诱导,形成NlABCB-dsRNA,喂养褐飞虱取食含NlABCB-dsRNA人工饲料,发现褐飞虱的存活率明显降低。
因此,褐飞虱NlABCB基因的表达特征表明,利用农药防治褐飞虱的最佳时期是在早龄期。这些结果可能有利于开发一些以NlABCB基因为靶标的安全有效的杀虫剂,同时可以利用RNA干扰的方法来抑制ABC转运蛋白的表达,达到控制害虫的目的。
