铜胁迫下两种生态型杠板归形态生理的差异性
2020-05-07张文玉
吴 曼,龚 云,彭 颖,张文玉,邹 添,陈 亮
(1.湖北师范大学 生命科学学院,湖北 黄石 435002; 2.湖北大别山国家级自然保护区英山管理局,湖北 黄冈 438700)
0 前言
铜是植物生长发育必需的微量营养元素, 在细胞中,铜元素的存在形式为Cu+和 Cu2+,在植物的生长发育过程中行使重要功能,以含铜蛋白的形式参与各种新陈代谢过程,如光合作用、呼吸作用电子传递链、活性氧代谢、对乙烯的感受、氮代谢、细胞壁的木质化及花粉的形成等[1]。植物在根毛部位协同水分代谢活动,能够通过单向运输载体蛋白顺着浓度梯度单向运输或主动运输把铜运至细胞内。通过共质体、质外体途径运输到根的内部,再由蒸腾拉力和根压运输到植物的各个部位,以酶的方式参与植物体内的氧化还原反应,并影响植物的呼吸作用。但是当环境条件中铜的含量超过其最大承受时,植物就会受到胁迫,进而对植物造成影响。
黄石大冶铜录山铜矿遗址土壤中累积大量的重金属铜矿废弃物,造成严重的土壤重金属污染[2~3],但杠板归的生长却十分繁茂。杠板归(PolygonumperfoliatumL),是一种一年生攀援草本植物,蔓生在野路旁。由于其中药功效,又名河白草、蛇倒退、梨头刺、蛇不过等。束文圣等人对该矿区杠板归等5种优势物种对重金属的吸收积累和迁移特性进行测定和比较,发现它们对Cu表现出了很高耐性[2]。且许多研究表明,杠板归对锰等重金属具有较强的富集作用,可以用于锰等重金属污染的治理修复[4]。但刘小红等人发现虽然杠板归以及鸭趾草对长江沿线铜矿区Cu有较多的吸收[5],却不足以达到富集量与生物量比值大于1000 mg·kg-1的超富集水平[6]。尽管目前有关杠板归的研究工作已经取得一定进展,但是大部分研究主要集中在将杠板归作为中药材论述其应用以及栽培技术等方面,在对铜的抗胁迫作用的研究却鲜见报道。我们在铜录山铜矿区和生活区(湖北师范大学区划范围内)都发现杠板归的分布,但在矿区的耐受型和生活区的敏感型杠板归植株颜色和叶子形状都有明显差异。由此我们实验室采集矿区耐受型杠板归和对照组敏感型杠板归,通过对比铜离子对两种生态型杠板归形态和光合作用两方面生理特征的影响,希望可以初步探究杠板归抗铜离子胁迫的生理机理,并为矿区对铜等重金属的生物治理提供理论支撑和指导。
1 实验方法
1.1 杠板归样本采集与培养
采集来自大冶市铜录山和湖北师范大学校园内的杠板归,采样时尽量选取生长良好的幼嫩枝条,从顶端往下剪取10~15cm枝条。将采集到的杠板归进行水培,用CuSO4·5H2O以1/2Hoagland营养液作为基础液配成不同浓度梯度的含铜离子的培养液,分别为0 μg/mL,100 μg/mL,200 μg/mL,300 μg/mL,400 μg/mL 5个浓度梯度,每个梯度每种类型2个重复,置于正常光照下培养。培养过程中每隔3天换一次培养液,培养约7天。
1.2 形态特征的测定
每组选取3~5株材料长势一致、生长状态良好的杠板归进行根长、株高等形态指标测定。测量根长(即第一个根毛基处至最末端的根尖之间的长度)和茎长(即由第一个根毛基处至茎顶端的长度)。
1.3 叶绿素含量的测定
从每个浓度铜离子处理的杠板归中选取叶片0.5 g,剪成碎块,每组设置3个重复,利用丙酮提取法浸提,以80%丙酮为对照,分别测定663 nm、645 nm处的吸光值[7]。根据Lamber-beer定律,利用公式:
Ca=12.7OD663-2.69OD645Cb=22.9OD645-4.68OD663
CT=Ca+Cb
求出红、绿杆马齿苋中叶绿素a、b的含量。
1.4 光合指标的测定
从每个浓度铜处理的杠板归选取6片大且无损伤的叶片利用 TPS-2光合仪进行叶片蒸腾速率(E),气孔导度(G),净光合速率(A),细胞间CO2的浓度(CI)的测量。
2 实验结果
2.1 形态特征比较
2.1.1 两种杠板归根茎叶形态差异
通过形态上观察,我们发现耐受型(铜录山组)根系较为发达,基部粗壮,茎干直径较大;敏感型(校园本地)根部较弱,且茎干直径较小植株多呈现倒伏状。如图1所示。
A B

2.1.2 茎长和根长的增长情况
由图2分析得,两种生态型的杠板归的根与茎的增长率分别呈先上升后下降的趋势,且耐受型的根和茎的增长率一直高于敏感型的杠板归。同时,敏感型的根与茎在100 mg/L铜离子浓度时有最大增长率,耐受型的根与茎在150 mg/L的铜离子浓度时有最大增长率,且耐受型杠板归植株的茎秆和根系增长率呈现较大的区别。

图2 不同Cu2+浓度下两种生态型杠板归的根和茎的增长率,不同字母表示在0.05水平的显著差异
2.2 光合指标比较
由图3可知,在经过不同铜浓度处理两周后,敏感型杠板归的气孔导度、蒸腾速率均随着铜离子的浓度增高呈减小的趋势,且铜离子浓度为200 mg/L时,其数值最小;而耐受型杠板归的气孔导度、蒸腾速率均随着铜离子浓度的增高呈增大的趋势,且铜浓度为150 mg/L时,其数值最大(A、C图)。
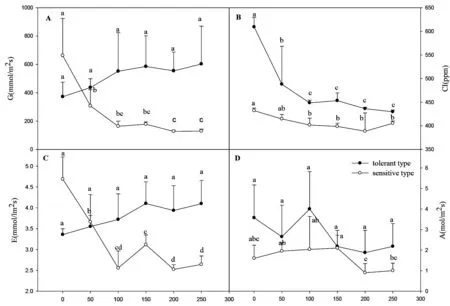
Cu2+Concentration(mg/L)
图3 不同铜离子浓度处理下两种生态型杠板归气孔导度、胞间二氧化碳浓度、蒸腾速率、光合速率指标
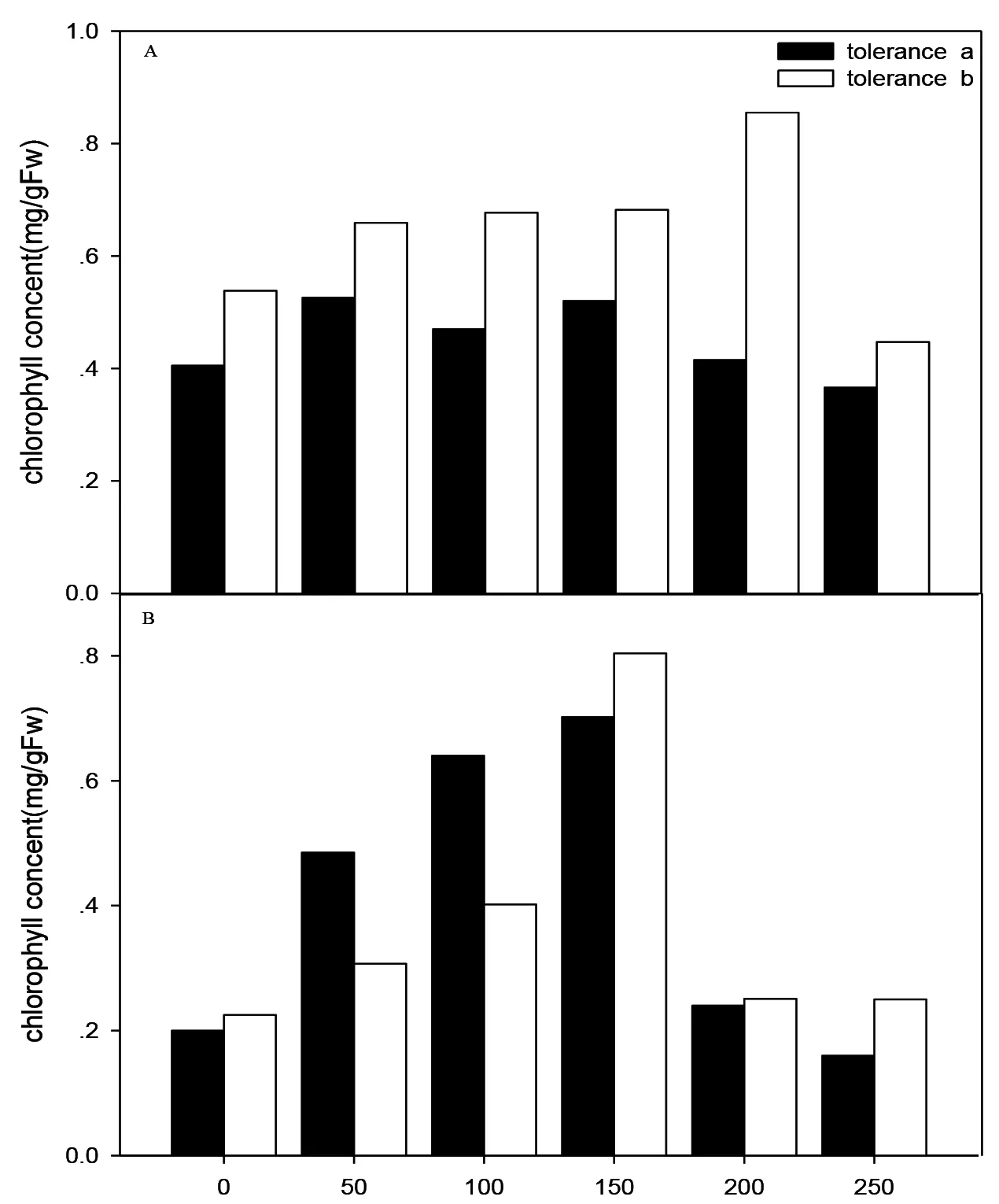
Cu2+Concentration(mg/L)
图4 不同铜离子浓度下杠板归叶绿素总含量
耐受型和敏感型杠板归的细胞间二氧化碳浓度、光合速率均随着铜浓度的增加而降低,且耐受型杠板归在铜离子浓度为200mg/L时,光合速率最低为1.88;在铜离子浓度为100 mg/L时最高达到4.00;敏感型杠板归在铜离子浓度为200 mg/L时,光合速率最低为0.90;在铜离子浓度为150 mg/L时最高达到2.10 .但耐受型杠板归胞间二氧化碳浓度、光合速率始终在敏感型杠板归之上(B、D图)。
2.3 叶绿素含量比较
由图4 A可知,耐受型杠板归的叶绿素b的含量要高于叶绿素a的含量,叶绿素b的含量在铜离子浓度为200 mg/L时达到最大,叶绿素a的含量在铜离子浓度为150 mg/L时达到最大;由图4 B可知,敏感型杠板归的叶绿素a的含量首先高于叶绿素b的含量,在铜离子浓度为150 mg/L后,叶绿素b的含量要高于叶绿素a的含量,叶绿素b的含量在铜离子浓度为150 mg/L时达到最大,叶绿素a铜离子浓度为150 mg/L时达到最大。
3 讨论分析
植物常常通过在形态和生理等方面采取不同的响应机制来应对不良环境。目前,国内外学者研究了多种植物在铜污染条件下的生长,结果表明,低浓度的铜离子能够促进植物种子萌发和生长,但当铜离子浓度继续增加后,对植物生长产生不同程度的抑制作用[8~9],甚至死亡[10]。如张刚等人[11]研究发现适量的铜对黑麦草生长起到促进作用,但是超出了阈值的铜对生长有抑制,会使其生长缓慢,根停止生长,最终枯死;植株矮化,叶黄化,这与我们的实验结果一致(图1),验证了我们的结论。另外,由根和茎的增长率可以看出(图2),杠板归在适应不同的环境时,会产生不同的响应机制,在营养分配上的表现较为突出。在离子胁迫下,刺激根系生长,改变生长中心,调控生长优势部位,减低地上蒸腾部位比例,以适应环境变化。而在我们实验中耐受型较之于敏感型发达的根系以及更强的根茎增长率也进一步验证了这个论断。
光合作用是植物生命活动的基础,是产生能量的重要来源,因此环境胁迫下植物光合作用的强弱也是反映植物生长状况重要生理指标。王钧[12]等人研究表明与对照组相比,在低浓度处理时耐受型杠板归的气孔导度、光合速率均上升,胞间二氧化碳浓度下降,高浓度处理后两种杠板归的气孔导度、光合速率均下降,胞间二氧化碳浓度上升。本实验中,随着铜离子浓度的升高,耐受型杠板归的各项指标始终位于敏感型之上,通过文献调研我们知道高等植物进行光合作用第一重要生产的物质是叶绿素,它含量的高低是影响光合作用强弱的重要因素,因此叶绿素动态变化可以反映植物受环境胁迫后的生理状态。研究发现,在Cu2+胁迫处理下,活性氧在细胞中增加,其更多地扩散到叶绿体内,从而加速了叶绿素的降解[13];同时,由于Cu2+局部积累过多,与叶绿体中蛋白质-SH 结合或取代其中的Fe2+、Zn2+、Mg2+,致使叶绿素蛋白中心离子组成发生变化而失活[14],且叶绿素a不及叶绿素b稳定,所以叶绿素a的降幅大于叶绿素b 。本试验结果表明,不同浓度Cu2+处理下,敏感型杠板归叶片叶绿素总量、叶绿素a、b含量明显变化,且叶绿素a / b下降;耐受型杠板归叶片叶绿素总量、叶绿素a、b含量变化较为平缓,且叶绿素a / b比值增大,由此说明在同等胁迫条件下铜录山(耐受型)杠板归光合作用相对于敏感型更强,因而植株更为健壮从而增强对胁迫的耐受型。
当然除开外观形态、光合生理指标方面,耐受型和敏感型植株都表现出明显差异,且通过我们实验也证实在较高浓度铜胁迫条件下植株生长更为良好,但是这两种不同生态型杠板归在铜胁迫下,其他生理指标中存在哪些差异仍需进一步探究。
