葡萄酒酿造过程中谷胱甘肽的研究进展
2020-04-25苏静,龚荣
苏 静,龚 荣
(山西农业大学食品科学与工程学院,山西 晋中 030801)
近年来,随着人们对谷胱甘肽(glutathione,GSH)认识的不断深入,研究者发现这种广泛分布于自然界的小分子肽与人类生产和生活有着密切关系。GSH是葡萄果肉中天然存在的小分子巯基化合物,是一种内源性抗氧化物质。葡萄酒酿造期间,GSH能够有效防止葡萄酒中酚类物质的氧化,保护葡萄酒的芳香物质和色泽;在葡萄酒贮藏阶段,GSH能够防止葡萄酒变质,并延长酒的货架期。2015年第38届世界葡萄与葡萄酒大会暨第13届国际葡萄与葡萄酒组织大会通过了两项决议,分别是向葡萄汁(OIV-OENO 445—2015)和葡萄酒(OIVOENO 446—2015)中添加还原型GSH的操作规范,建议在葡萄汁和葡萄酒中添加还原型GSH来保护葡萄酒的色泽和风味,规定最大使用剂量均为20 mg/L。然而,这种做法并没有获得欧盟的认可。另外,许多葡萄酒生产厂商也选择采用高产GSH的酿酒酵母(Saccharomyces cerevisiae)或者非活性干酵母(又称为惰性酵母或灭活酵母)来增加酒中还原型GSH的含量,以改善葡萄酒品质。
近年来,国外关于葡萄、葡萄汁和葡萄酒酿造过程中还原型GSH作用的研究较多,而国内研究较少。这些研究大多数集中在葡萄中GSH或者果香硫醇前体物对葡萄酒挥发性硫醇的影响,GSH对葡萄酒中化学成分尤其是香气物质和色泽的影响;葡萄酒酿造过程中GSH含量的变化;高产GSH酵母菌株的筛选及其对乙醇发酵的影响等方面。全面认识和理解GSH在葡萄酒酿造过程中的作用,对更好地监控并提高GSH含量,防止葡萄酒氧化及保护葡萄酒的香气具有重要意义,这也为高品质葡萄酒的酿造提供一定的理论基础。本文分析了国内外相关文献,以对葡萄酒酿造过程中GSH的研究进展进行综述。
1 谷胱甘肽的结构和特性
GSH是由谷氨酸、半胱氨酸、甘氨酸三种氨基酸组成的三肽,是生物体中(包括植物、动物和真菌)最普遍存在的细胞内巯基物质,具有很强的抗氧化性。GSH中的半胱氨酸残基(巯基部分)赋予其独特的氧化还原反应和亲核性能,使其成为生物体抗氧化系统中重要的组成成分。GSH具有两种存在形式,分别是还原型GSH和GSH二硫化物(glutathione disulfide,GSSG)[1],见图1、2。

图1 还原型GSH(A)和GSSG(B)分子结构[2]Fig. 1 Molecular structures of reduced GSH (A) and glutathione disulfide (B)[2]
当机体发生氧化应激时,还原型GSH和GSSG之间可以进行相互转换。还原型GSH能够在GSH过氧化物酶(glutathione peroxidase,GSH-Px)催化下转化为GSSG和水,而GSSG也能够在GSH还原酶(glutathione reductase,GR)作用下,以NADPH作为辅因子转化为还原型GSH。此外,还原型GSH还参与了氧化蛋白的还原。谷氧还蛋白(glutaredoxin,Grx)是一种修复蛋白质活性的小分子酶蛋白,在GSSG与还原型GSH之间进行转化的过程中,Grx中的二硫键被还原为巯基[3-4]。
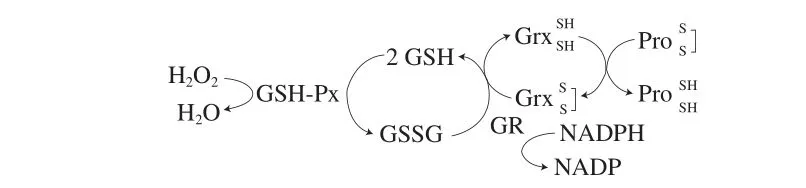
图2 还原型GSH在细胞中的主要功能[3--4]Fig. 2 Major functions of reduced GSH in cells[3-4]
2 葡萄果实中的谷胱甘肽
1989年,Cheynier等[5]首次在葡萄中发现了GSH。葡萄浆果中90%以上的GSH都是还原型GSH,其含量为17~114 mg/kg[5-10]。在外界条件良好的情况下,植物叶片中还原型GSH和GSSG的比例约为20∶1。随着葡萄的成熟,葡萄浆果中还原型GSH的含量逐渐增加。葡萄品种、葡萄采收期和葡萄栽培技术等都会影响还原型GSH含量。研究发现,降低叶片中还原型GSH含量可以增加葡萄浆果中还原型GSH的含量,因此,可以在葡萄浆果刚开始进入成熟期时就进行栽培管理,使叶片中的还原型GSH转移到葡萄藤的韧皮部位,继而转移到葡萄浆果中;此外,摘叶处理也会增加葡萄中还原型GSH的含量,然而这一现象并不具有普遍性[7,10-11]。还原型GSH在葡萄中积累的机制以及不同的栽培方式对葡萄中还原型GSH含量的影响仍是今后研究的重点。
还原型GSH还与某些品种葡萄香气的形成有关。‘长相思’是深受人们喜爱的白葡萄品种,虽然其果香不浓郁,但其所酿造的葡萄酒却具有独特且浓郁的风味。这与果实中含有丰富的硫醇类物质有关,这类物质赋予‘长相思’葡萄有别于其他葡萄品种独特的水果香气,如葡萄柚、柑橘、百香果等热带水果的香气[8]。葡萄果实中的还原型GSH可与硫醇结合而形成结合态硫醇前体物[8]。‘长相思’葡萄中重要的硫醇前体物为GSH类和半胱氨酸类前体化合物,包括S-3-(正己醛)-GSH、S-3-(正己醇)-GSH(S-3-(hexan-1-ol)-glutathione,GSH-3MH)、S-3-(正己醇)-L-半胱氨酸(S-3-(hexan-l-ol)-L-cysteine,Cys-3MH)、4,4-甲基异丁基酮-L-半胱氨酸(4,4-(methylpentan-2-one)-L-cysteine,Cys-4MMP)和4-S-谷胱甘酰-4-甲基异丁基酮(4-S-glutathionyl-4-methylpentan-2-one,GSH-4MMP)[12]。近年来,许多葡萄品种如‘雷司令’、‘赛美蓉’、‘梅乐’、‘赤霞珠’、‘歌海娜’中也检测到了这些果香硫醇前体物质[13-15]。
葡萄果实中的果香硫醇前体物质需要酿酒酵母的帮助才能够释放出挥发性香气物质。例如,酿酒酵母胱硫醚β-裂解酶可将胱硫醚类香气前体物的C-S键断裂,从而将果香硫醇类香气物质释放到葡萄酒中。有些研究者认为,GSH类前体物是通过脱除谷氨酸和甘氨酸而形成半胱氨酸偶联产物[16]。通常认为硫醇前体物的浓度应与挥发性硫醇的浓度有关,事实上,在发酵过程中只有一小部分的芳香硫醇被释放出来[17-19]。这与酿酒酵母中胱硫醚β-裂解酶的活性较低有关,据报道,只有3.2%的胱硫醚类香气前体物被裂解[20]。马捷等[20]构建了具有较高胱硫醚β-裂解酶活性的酵母工程菌株,其胱硫醚β-裂解酶活力与出发菌株相比提高了32.25%~59.44%,在‘长相思’干白酒中硫醇类物质的产量提高了1.22 倍。
3 葡萄汁中的谷胱甘肽
据报道,葡萄汁中还原型G S H质量浓度在1.1~102 mg/L之间[5,9,21-25],其含量与葡萄品种、葡萄汁中酪氨酸酶活性、氧气、浸渍和压榨等因素有关[7,9]。葡萄榨汁后,葡萄汁中还原型GSH含量迅速下降,它与对羟基肉桂酸乙酯发生氧化反应生成无色的反应产物——2-S-谷胱甘酰咖啡酰酒石酸,也被称为葡萄反应产物(grape reaction product,GRP)。白葡萄汁中GSSG质量浓度在0.46~2.93 mg/L之间,氧化造成葡萄汁中GSSG质量浓度上升,GSSG可以作为判断葡萄汁氧化程度的指示剂[7]。
‘长相思’葡萄压榨后,葡萄汁中Cys-3MH含量大幅增加,而GSH-3MH含量较少。这是因为在GSH S-转移酶作用下,GSH-3MH可被转化为Cys-3MH[26-27]。冻融处理也会影响葡萄中还原型GSH的代谢,从而造成葡萄汁中GSH-3MH含量的变化[12]。Thibon等[28]发现‘长相思’和‘赛美蓉’葡萄感染贵腐霉菌(Botrytis cinerea)后,其葡萄汁中挥发性硫醇前体物的浓度会大大提高。这是因为贵腐霉菌分泌的代谢物促进了(E)-2-乙烯醛的形成,它与还原型GSH结合后可转化为Cys-3MH[29]。
4 发酵和陈酿过程中的谷胱甘肽
4.1 谷胱甘肽与乙醇发酵
葡萄酒中的GSH一部分来源于葡萄本身,另一部分来自乙醇发酵阶段酵母菌的代谢。与葡萄和葡萄汁相比,葡萄酒中还原型GSH的质量浓度较低(0~70 mg/L),并且具有高度可变性[7,22,30-34]。葡萄品种、酿造工艺、酵母菌株、酒泥和陈酿环境等均会影响葡萄酒中GSH的含量。
在乙醇发酵过程中,葡萄汁中的还原型GSH成为酿酒酵母良好的“营养品”,它被酵母吸收并为酵母提供营养,促进了乙醇发酵进程。然而葡萄酒中还原型GSH含量的变化往往是比较复杂的,其含量可能会升高[21-22,24-25],这是因为酿酒酵母代谢并分泌出一部分还原型GSH或酵母吸收了还原型GSH前体物质合成了还原型GSH,从而导致发酵醪中还原型GSH含量增加。这一过程是由酿酒酵母细胞内Gex1基因进行调控[35-37]。酿酒酵母吸收和释放还原型GSH的程度取决于酵母菌株自身的遗传背景、代谢能力以及环境因素,如外界温度、pH值、渗透压、营养成分的含量和比例(碳源、氮源、半胱氨酸)等[21,38-39]。生长在富含高浓度还原型GSH环境中的酵母,其胞内还原型GSH含量也较高[21]。Park等[24-25]认为葡萄汁中总氮或可吸收氮的含量与葡萄酒中还原型GSH水平之间具有明显相关性,然而Kritzinger[30]对这一结果并不认可。乙醇发酵后还原型GSH含量还可能会下降[7,10,40]。Patel等[10]研究证实了这一过程,他们采用添加了高质量浓度(67 mg/L)还原型GSH的葡萄汁进行乙醇发酵,经检测,发酵后每升葡萄酒中仅有几毫克还原型GSH。这是因为酿酒酵母自身转运蛋白吸收了发酵醪中的还原型GSH或是由于还原型GSH转化成GRP所导致的[35,37]。
在原料和酿造工艺不变的条件下,酒中还原型GSH含量主要是由酿酒酵母代谢所决定。通过选育高产还原型GSH的酿酒酵母可以提高葡萄酒抗氧化的能力。目前,市场上存在一类富含还原型GSH的商业活性干酵母(GSH-inactive dry yeast,GSH-IDY),其具有提高葡萄酒中还原型GSH含量和增强葡萄酒感官稳定性的功能[30,41-43]。这类产品的生产厂商有拉曼(Lallemand)、拉氟德(Laffort)和英纳帝斯(Enartis)等。乙醇发酵过程中,GSH-IDY是通过释放自身还原型GSH和吸收还原型GSH前体物来增加葡萄酒中还原型GSH的含量。不同GSH-IDY释放还原型GSH水平不同,在1.45~2.53 mg/L之间,总GSH释放量在1.63~3.44 mg/L之间[21,30]。然而,并非所有GSH-IDY都能在乙醇发酵过程中释放还原型GSH,这可能与菌株、酿酒工业环境等因素有关[21]。此外,GSH-IDY通过释放甘氨酸、精氨酸、谷氨酸、蛋氨酸和半胱氨酸等多种氨基酸,不仅增加了葡萄酒中氨基酸的含量,也具有促进酿酒酵母产生还原型GSH的作用[39,41]。Kritzinger[30]认为具有较高氮吸收水平的GSH-IDY酿造的葡萄酒中还原型GSH含量也较高,并建议在乙醇发酵初期人工补加还原型GSH。此外,生产活性干酵母所需的热加工过程也可能造成还原型GSH释放量的改变。在GSH-IDY制备的干燥阶段,还原型GSH与还原糖之间发生美拉德反应会在一定程度上造成还原型GSH损失。许多研究者认为采用富含还原型GSH的酵母菌株进行葡萄酒乙醇发酵,能够显著提高成品葡萄酒中还原型GSH的含量[30,44-45]。然而,Fracassetti等[22]认为酵母菌株对葡萄酒中还原型GSH含量的影响甚微。乙醇、SO2等因素也会造成GSH-IDY释放还原型GSH量的差异[8],然而目前相关的报道较少。
筛选高产还原型GSH的酿酒酵母仍是目前研究热点之一[46]。酿酒酵母中还原型GSH的含量与菌株、生长环境等因素有关。硫酸盐还原途径是酿酒酵母细胞内含硫化合物合成的重要途径。酿酒酵母细胞内的GSH是γ-谷酰基-半胱氨酸、谷氨酸和半胱氨酸在γ-谷氨酰半胱氨酸合成酶(由GSH1基因编码)和GSH合成酶(由GSH2基因编码)催化下合成[47]。对钼酸盐具有高抵抗能力的酿酒酵母往往会分泌较多的还原型GSH。因此,研究者常利用钼酸盐进行高产还原型GSH酿酒酵母的筛选。其原因可能是钼酸盐激活了酿酒酵母中与金属胁迫响应相关的代谢途径,包括硫同化途径以及还原型GSH的合成和代谢途径[47]。Mezzetti等[47]采用基因组学和转录组学方法对这一相关机制进行研究,认为氨基酸通透酶、与氨基酸生物合成前体有关的酶以及与氧化胁迫有关的酶等也参与了这一过程。
将酵母灭活后获得的具有葡萄酒芳香保护能力的非活性干酵母产品也深受酿酒师喜欢。非活性干酵母富含还原型GSH和还原型GSH前体物质(如半胱氨酸),还含有酵母多糖(如甘露糖蛋白)、矿物质、微量元素等丰富的营养物质。在酒精发酵过程中,这类产品能为酵母提供营养,促进酿酒酵母中GSH前体物质的合成,从而进一步合成还原型GSH,保证酒精发酵的顺利进行。研究显示,在白葡萄酒发酵过程中,添加富含还原型GSH的非活性干酵母,能减少H2S的产生,稳定白葡萄酒的香气,改善葡萄酒的口感。
4.2 谷胱甘肽与苹果酸-乳酸发酵
苹果酸-乳酸发酵(malolactic fermentation,MLF)是将葡萄酒中二元苹果酸转化为一元乳酸和二氧化碳的过程,从而降低葡萄酒酸度,提高葡萄酒的稳定性,并丰富葡萄酒风味、增强酒的结构感和馥郁度。酒酒球菌(Oenococcus oeni)是MLF的主要启动者,然而葡萄酒中高浓度的乙醇、较低的pH值以及高SO2含量等因素,往往使这一发酵过程变得缓慢,甚至难以启动。研究表明,MLF过程中,葡萄酒中总GSH含量显著降低,降低量约为初始值的21%~36%;而还原型GSH和GSSG的变化并没有明显规律[48]。
还原型GSH能够提高酒酒球菌对酸胁迫或/和乙醇胁迫环境的耐受性,增强其生长能力,提高MLF速率[3-4,49-51]。虽然酒酒球菌不能合成还原型GSH,但它具有吸收外源还原型GSH的能力,这种能力因菌株的不同而有差异[4]。酒酒球菌中含有一些与还原型GSH代谢相关的酶和蛋白,包括Grx、GSH-Px、GR和假想还原型GSH转运蛋白[52-55]。还原型GSH提高酒酒球菌环境胁迫耐受性的机理可能是调节其细胞膜脂肪酸成分以适应环境,并且提高了菌株胁迫适应性相关基因hsp18和clp在发酵初期的相对表达水平[4]。谷氧还蛋白基因grx和cydC是参与还原型GSH保护酒酒球菌抵御酸或乙醇胁迫过程中的关键基因[49]。还原型GSH(5 mmol/L)的添加使得供试菌株中基因rdx3(OEOE_RS04215)、cydC和rdx2高表达[49]。苏静[51]在添加了还原型GSH的培养基中对酒酒球菌SD-2a进行预适应性培养,发现与酒酒球菌胁迫适应性相关的基因hsp18、citE、clpP、ctsR和rmlB在MLF前期的相对表达水平提高,并建议采用添加还原型GSH的培养基对菌株进行预培养,以此来提高其在葡萄酒中的生存能力并促进MLF进程。另外,GR在酒酒球菌处理由乙醇胁迫导致的氧化损伤和维持细胞氧化还原平衡方面发挥重要作用[50]。
虽然研究发现还原型GSH能够增强酒酒球菌的酸或/和乙醇胁迫耐受性,但是其机理并不十分清楚。酸或/和乙醇应激反应过程是一个非常复杂的网络体系,添加的外源氨基酸可能通过诱导菌体内部其他蛋白质发生变化来应对这些胁迫。关于还原型GSH、MLF以及酒酒球菌之间的关系,以及如何更好地利用还原型GSH来促进MLF进程,提高葡萄酒质量,未来还需要进一步的探索。
4.3 谷胱甘肽与陈酿
无论何种类型的葡萄酒,陈酿后酒中还原型GSH含量均逐渐降低[11,21,30,45],且其下降幅度大于总GSH的下降幅度[30]。葡萄酒进行压榨操作,不仅促进了多酚的氧化,还会造成还原型GSH含量快速下降,而陈酿后酒中还原型GSH含量的下降幅度更大[10-11]。Lavigne等[45]也发现,经压榨后的葡萄酒若在新橡木桶中陈酿,会因木材的氧化反应导致消耗还原型GSH的速率加快。
白葡萄酒对氧气非常敏感,陈酿时要特别注意酒中氧气的浓度。通常葡萄酒在低氧环境中受到氧化的程度较低,陈酿后的酒中还原型GSH含量较高。Arapitsas等[56]发现半胱氨酸和还原型GSH在葡萄酒陈酿期间会发生磺化反应形成半胱氨酸硫酸盐和GSH硫酸盐。他们认为这与葡萄酒装瓶时接触了大量的氧气有关。此外,Lavigne等[45]认为酒泥可能具有保护还原型GSH的作用。
5 谷胱甘肽在葡萄酒酿造过程中的作用
5.1 谷胱甘肽防止葡萄汁和葡萄酒的氧化并保护葡萄酒的色泽
葡萄酒酿造过程的实质是酵母菌、乳酸菌等微生物进行生物转化的过程,这个过程易受到环境温度、pH值、氧以及不良微生物等的影响。葡萄浆果破碎后迅速接触到大量氧气,在多酚氧化酶(polyphenol oxidase,PPO)催化下发生酶促氧化褐变,葡萄汁中的酚类物质被氧化成邻苯醌,邻苯醌通过与葡萄汁中的活跃成分进行聚合和缩合形成褐色物质,致使葡萄酒氧化褐变[57]。Li Hua等[57]认为在葡萄酒众多多酚类物质中,咖啡酸和儿茶素是最易被氧化的邻苯二酚类物质。在葡萄酒陈酿阶段,这些物质还可经非酶氧化(化学氧化)形成邻苯醌。传统葡萄酒工业生产中常采用添加SO2的方式防止葡萄汁和葡萄酒的氧化褐变,SO2通过不可逆地与PPO(氧化型和过渡型)发生络合作用,从而抑制PPO活性。这种方法不仅廉价而且有效,因此一直沿用至今。然而,过量使用SO2会使葡萄酒色泽异常,还会造成人体过敏,危害人体健康。
还原型GSH能够防止葡萄汁和葡萄酒的氧化褐变[11,58-67]。虽然还原型GSH不能抑制PPO活性,但其可以通过与醌式咖啡酰酒石酸(已被氧化的酚类物质)反应形成GRP,从而抑制邻苯醌的形成和褐色聚合物的产生[8]。此外,当还原型GSH浓度达到一定值时还可诱捕邻苯醌,限制邻苯醌引发的其他反应[68-71]。研究人员采用超高分辨率质谱代谢组学和感官分析相结合的方法证实了还原型GSH在白葡萄酒陈酿期间发挥了保护性作用,通过保护葡萄酒中的含氮、含硫物质、氨基酸、芳香物质、多肽等抗氧化代谢物来发挥其保护作用[72]。然而,SO2和铜离子等物质也会在一定程度上限制其抗氧化作用的发挥。SO2与儿茶酚酶相互作用,生成游离咖啡酰酒石酸和对香豆酰酒石酸,而GRP容易被漆酶和醌式咖啡酰酒石酸氧化褐变[73]。铜离子可与醌式咖啡酰酒石酸相互竞争争夺还原型GSH,因而降低了葡萄酒中还原型GSH的有效浓度,限制了GRP的生成。此外,醌式咖啡酰酒石酸与黄烷醇的偶合氧化也增加了葡萄汁褐变的程度[5,69]。
花色素苷是构成红葡萄酒颜色的一类重要的多酚类物质。红葡萄酒颜色从红色到黄色的转变是葡萄酒老化的标志,也是葡萄酒氧化变质的表现。葡萄酒的氧化分为两种:第1种是上述提到的形成醌类物质,O2被还原产生了H2O2;第2种是酒中存在铁和铜离子等微量金属离子时,发生了芬顿(Fenton)反应,产生羟自由基。二甲花翠素-3-葡萄糖苷是葡萄酒中重要的花色素苷,芬顿反应产生的羟自由基是造成其降解的主要原因。研究证实,在pH值(pH 3.2)较低的模拟葡萄酒和红葡萄酒中,SO2能够清除酒中存在的H2O2,并与因芬顿反应而产生的羰基化合物结合,是保护酒中二甲花翠素-3-葡萄糖苷最有效的抗氧化剂[74]。然而,单独使用还原型GSH或者与SO2结合使用时,还原型GSH均不能起到清除H2O2的作用,因而不能抑制芬顿反应,也不能防止二甲花翠素-3-葡萄糖苷的降解。这是由于在H2O2存在的氧化环境中,还原型GSH转变为GSSG,GSSG与磺酸根离子(SO3H-)反应产生磺化GSH,从而造成还原型GSH的大量减少[75]。因此,葡萄酒中还原型GSH发挥抗氧化作用的机理主要是通过限制醌类物质的合成来防止褐变,并不能通过清除酒中存在的H2O2来抑制芬顿反应。
还原型GSH防止葡萄酒氧化褐变以及稳定陈年葡萄酒色泽的效果依赖于其使用剂量[59-60,66,76]。研究人员在干白葡萄酒进行瓶储前,向酒中加入10 mg/L的还原型GSH,结果表明还原型GSH能够有效抑制陈酿过程中葡萄酒的酶促褐变,从而防止葡萄酒色泽变黄,并减少酒中异味的生成[58]。Dubourdieu等[60]也发现类似的现象,他们认为如果陈酿前白葡萄酒中还原型GSH质量浓度超过6~10 mg/L,成品酒的颜色就可以得到更好的保护。梁晓芳等[77]向龙眼干白葡萄酒中添加20 mg/L还原型GSH,有效地减缓了酒中酚类物质的减少和氧化褐变,延长了葡萄酒的存放时间。然而,也有研究发现,还原型GSH有可能会破坏葡萄酒的色泽,这可能与使用的测定体系有关。Sonni等[78-79]将模拟葡萄酒贮存在温度为45 ℃、氧气充足且无SO2的环境中,此时,还原型GSH能够抑制酒中o-醌衍生物酚类化合物和黄色呫吨鎓阳离子色素的形成;单独添加还原型GSH时,葡萄酒中形成一些与对照酒(不添加还原型GSH)中聚合色素结构不同的新的聚合色素,而这种新的聚合色素有可能会对葡萄酒的色泽产生影响;该研究还发现复合使用还原型GSH(860 mg/L)和VC(500 mg/L)能够更好地保护葡萄酒的色泽,当酒中存在VC时,高质量浓度的还原型GSH(860 mg/L)能够延缓VC的降解,抑制VC降解产物与黄烷醇、(+)-儿茶酸反应,且未形成上述新的聚合色素。然而,当VC耗尽,还原型GSH又会造成酒中聚合色素的生成。
5.2 谷胱甘肽保护葡萄酒的风味
香气是衡量葡萄酒品质的重要指标之一。葡萄酒的香气来源于葡萄本身以及酿造和成熟过程中丰富的芳香物质,包括酯类、醇类、醛类、萜烯类、硫醇类、酮类等[8]。其中,高级醇、乙基酯类和萜烯醇类物质能够赋予葡萄酒果香味。2-苯乙醇是一种具有玫瑰香味的芳香醇,乙酸苯甲酯具有苹果香气,乙酸异戊酯具有香蕉的气味,芳樟醇、萜品醇、橙花醇、香叶醇、脱氢芳香醇赋予葡萄果香、似玫瑰的香味[8]。葡萄酒中酯类、萜烯类、醇类物质含量的下降意味着葡萄酒风味的损失。在葡萄酒酿造过程中,葡萄酒风味的变化可能是由于酒中芳香类物质发生了氧化或其他化学反应,如酯类物质发生酯化和水解反应,萜烯醇转化为萜类氧化物,芳樟醇转化为具有更高感官阈值的萜品醇等[80-81]。葡萄酒中的芳香物质还会由于外界环境的变化如空气氧化、温度、化学反应等而导致香气成分流失,甚至产生异味。
许多研究证实,还原型GSH可以使葡萄酒更好地保留某些重要的酯类、萜烯类和硫醇类物质[8],防止其损失。研究发现,在白葡萄酒进行陈酿前添加还原型GSH,可以更好地保留乙酸异戊酯、己酸乙酯、辛酸乙酯、癸酸乙酯、芳樟醇和里那醇等重要的香气物质[61,63-65]。还原型GSH保护葡萄酒芳香物质的机制可能是由于其结构中存在游离的巯基组分,因而使其具有很好的自由基和过氧化物清除能力。还原型GSH对葡萄酒香气物质的保护效果与其质量浓度有关[61]。Dubourdieu等[60]认为白葡萄酒中还原型GSH的添加量大于6~1 0 mg/L,可以起到保护陈酿葡萄酒香气的作用。Papadopoulou等[61]研究表明,在葡萄酒陈酿前,向酒中添加20 mg/L还原型GSH能够防止该葡萄酒中芳樟醇和萜品醇含量的下降。Roussis等[64]比较了单独使用SO2(50 mg/L)以及SO2(35 mg/L)与还原型GSH(20 mg/L)复合使用时对瓶储后葡萄酒香气物质的影响,发现后者能够更有效地保护葡萄酒中的酯类物质和芳樟醇,并且有效地保护酒中的酚类物质尤其是咖啡酸。此外,Webber等[82]发现还原型GSH的添加对葡萄酒中游离SO2、总酚、儿茶素、表儿茶素、咖啡酸和香豆酸的影响很小或者几乎没有影响,并且保护了陈酿后葡萄酒中香气的复杂性和新鲜度。
硫醇类物质是葡萄酒中重要的香气成分。在葡萄酒进行瓶储期间,低浓度的还原型GSH对酒中的硫醇类物质具有保护作用。‘长相思’白葡萄酒具有浓郁的水果香气,这主要归功于其中重要的硫醇类物质,包括4-巯基-4-甲基-2-戊酮(4-mercapto-4-methylpentan-2-one,4MMP)、3-巯基-1-己醇(3-mercaptohexan-1-ol,3MH)和3-巯基己基乙酸酯(3-mercaptohexyl acetate,3MHA),这些物质具有黄杨木、西番莲、葡萄柚和黑醋栗等水果的味道[83-86]。其中,3MH和Cys-3MH分别是由Cys-3MH和GSH-3MH产生的,4MMP是由Cys-4MMP和GSH-4MMP产生的[8,26,87]。乙醇发酵期间,在酿酒酵母的作用下,葡萄酒中生成了葡萄中所没有的4MMP和3MH这两种挥发性的硫醇。然而,在葡萄酒瓶储期间,这些挥发性硫醇物质逐渐消失,这是因为硫醇类物质极易受到氧气的影响[8,30],一般与多酚氧化、金属催化氧化有关。研究表明,在‘长相思’葡萄酒进行瓶储前,向酒中添加10 mg/L或20 mg/L的还原型GSH均能显著提高陈酿后葡萄酒中3MH的含量[11,88]。同时添加SO2与还原型GSH也可以减缓‘长相思’葡萄酒中3MH和3MHA的降解[59]。此外,葡萄酒中的羟基肉桂酸易被氧化形成o-醌,这些醌易与硫醇发生迈克尔加成反应[8]。而还原型GSH含有的巯基组分也能与o-醌发生反应,因此,它与硫醇之间进行竞争,从而防止硫醇类物质在葡萄酒瓶储期间遭受破坏[89]。然而,Patel等[10]采用含高质量浓度还原型GSH(67 mg/L)的葡萄汁进行发酵,发酵后的酒中多种硫醇类物质含量下降,然而其下降机理并未得到解释。
20世纪80年代末期,研究人员在德国葡萄酒中发现了两种能导致葡萄酒贮存时产生异味的物质,分别是葫芦巴内酯(3-羟基-4,5-二甲基-2(5H)-呋喃酮)和2-氨基苯乙酮(2-aminoacetophenone,2-AAP)[60]。前者在葡萄酒中的感官阈值为8 μg/L,具有强烈的咖喱味;后者在葡萄酒中的感官阈值为0.5~1.5 μg/L,若其质量浓度高于感官阈值,会呈现湿羊毛、杂醇油、臭樟脑和家具擦光油的味道[90]。Dubourdieu等[60]在‘长相思’葡萄酒装瓶前添加10 mg/L还原型GSH,将其贮存3 年后,发现酒中葫芦巴内酯含量在其感官阈值以下,酒中2-AAP含量为对照葡萄酒中的一半,从而证实了还原型GSH具有抑制这两种物质生成的作用,然而其作用机制并未得到阐明。
H2S是葡萄酒中具有臭鸡蛋味的物质,当其在葡萄酒中的含量超过其感官阈值时就会严重影响葡萄酒的品质。葡萄酒中H2S的含量受到葡萄醪中含硫化合物含量、酵母菌株、发酵条件、葡萄汁营养状态等因素的影响[8]。研究发现,还原型GSH与H2S的产生有关,还原型GSH中的半胱氨酸残基可在半胱氨酸脱巯基酶的作用下形成H2S。葡萄酒含有高浓度的还原型GSH或者处于低氧环境且含有二价铜离子的环境中,都会促进H2S的产生[11]。Kreitman等[91]发现,铜离子与还原型GSH、H2S、甲硫醇(methanethiol,MeSH)共同存在时会产生MeSH-GSH二硫化物和MeSH-GSH三硫化物。因此,葡萄酒中含有高浓度的还原型GSH会对葡萄酒的风味产生不利影响。目前,关于还原型GSH利于H2S产生的机理仍未得到充分阐明。针对不同葡萄原料和酿造工艺,研究者应寻找还原型GSH的最佳添加剂量和时间,并控制H2S生成量,扬长避短,以达到提高葡萄酒品质的目的。
6 结 语
目前,GSH已经得到了各国学者和酿酒师的普遍关注。GSH是葡萄和葡萄酒的内源性抗氧化剂,也是一种安全有效的葡萄酒品质改良剂,在保护葡萄酒色泽和风味的同时增加了葡萄酒的营养。此外,提高葡萄酒内源性还原型GSH的含量可以减少SO2的使用量,有效抑制葡萄酒氧化,避免因使用大量SO2而造成葡萄酒风味协调性差以及产生不良异味的现象。目前,可以通过以下措施来提高葡萄和葡萄酒中还原型GSH的含量:在葡萄种植期间,采用合理的栽培措施增加葡萄果实中还原型GSH的含量;在葡萄酒酿造过程中,选择在葡萄汁中添加还原型GSH、使用非活性干酵母或采用高产还原型GSH酿酒酵母进行乙醇发酵;在葡萄酒瓶储前,采取添加外源食品级还原型GSH的方法防止葡萄酒氧化和香气丧失。然而,酒中含有高浓度的还原型GSH可能会促进具有不良气味H2S的产生。因此,针对不同的葡萄品种、葡萄酒类型以及酿造工艺,寻找适合的还原型GSH添加时间和剂量非常关键,同时未来需综合运用各种分析技术进一步探索还原型GSH如何影响葡萄酒的品质。如研究还原型GSH在乙醇发酵、MLF、贮藏过程中的变化规律;外源还原型GSH的添加时间和添加剂量对葡萄酒品质的影响;高产还原型GSH酿酒酵母对葡萄酒品质的影响;还原型GSH对葡萄酒中酚类、芳香类物质、酵母菌及乳酸菌的影响及其作用机理;影响葡萄酒中GSH含量的因素等。这些相关研究将为还原型GSH应用于葡萄酒的酿造提供更详实的参考和实践支持。
