矮牵牛同源三倍体种质创制及减数分裂观察
2020-04-17魏跃陈景景郑宏伟许建颜志明孙朋朋
魏跃 陈景景 郑宏伟 许建 颜志明 孙朋朋


摘要:为创制矮牵牛三倍体种质并对其进行细胞学研究,以矮牵牛二倍体、四倍体为亲本进行正反杂交,对杂交后代进行染色体计数并与双亲进行农艺性状、解剖学比较,采用常规压片法对三倍体花粉母细胞减数分裂进行观察。结果表明:(1)以二倍体、四倍体为亲本进行正反杂交可获得三倍体矮牵牛,但结籽率均较低;(2)同源三倍体终变期大部分细胞内都出现单价体、二价体和三价体共存的现象,中期Ⅰ和中期Ⅱ出现游离在赤道板外的染色体,后期Ⅰ染色体有10/11、9/12、8/13和少数7/14分离方式,并出现姊妹染色单体提前发生分离和丢失染色体等异常现象。花粉母细胞减数分裂异常导致了大量不育花粉的形成,同源三倍体创制为初级三体系选育奠定了基础。
关键词:矮牵牛;同源三倍体;染色体;花粉;减数分裂
中图分类号: S681.601
文献标志码: A
文章编号:1002-1302(2020)02-0131-05
收稿日期:2018-12-19
作者简介:陈景景(1996—),女,江苏徐州人,主要从事园艺花卉育种与栽培研究。E-mail:695355008@qq.com。
通信作者:魏 跃,博士,教授,主要从事园艺花卉细胞学与遗传育种研究。E-mail:1069125418@qq.com。
同源三倍体为植株细胞具有三套来源相同的染色体组,具有广泛的应用价值,不但可用于杂种优势和无籽果实的生产[1],还可作为介质合成单体、三体等各种非整倍体遗传材料[2-3],其中单体、三体是进行基因定位、确定连锁群与染色体关系研究的重要遗传材料[4]。同源三倍体一般是通过二倍体、四倍体杂交获得,在橙子[5]、西瓜[6]、甜瓜[6]、葡萄[7]、結球甘蓝[8]等作物上都已有成功报道。
矮牵牛(Petunia hybrida Vilm.)为茄科矮牵牛属1年生草本花卉,是目前广泛应用的重要花坛花卉之一。近年来,国内研究人员已对矮牵牛开展了聚类分析[9]、花药培养[10]、基因克隆[11-12]、遗传转化[13]和多倍体[14]等多方面研究,但关于三倍体种质创制及相关细胞学研究还没有过报道。
本研究以二倍体、四倍体矮牵牛为材料进行同源三倍体矮牵牛种质创制,并对其花粉母细胞减数分裂进行观察和分析,这不仅丰富了矮牵牛种质资源创制及相关细胞学内容,也为系列初级三体等非整倍体创制和细胞遗传学研究奠定基础。
1 材料与方法
1.1 试验材料
同源四倍体材料07-12为江苏农林职业技术学院花卉遗传与种质创新课题组培育,二倍体品种梦幻(蓝色)来源于浙江虹越花卉股份有限公司,后经过多代自交保存。
1.2 试验方法
1.2.1 同源三倍体的获得与鉴定 2015—2016年3月在江苏农林职业技术学院园艺试验基地(119.17°E、31.95°N)播种07-12和梦幻,6月上旬2个亲本进入花期后以07-12、梦幻分别为父母本进行正反杂交,每次分别授粉50朵,重复3次,7~10 d后统计坐果率,23~35 d后统计单果结籽数,同时2个亲本分别自交作为对照。坐果率=果实数/授粉花朵数×100%;单果结籽数=饱满种子数/果实数。
2016—2017年3月播种矮牵牛F1杂交种子,5月中旬F1植株进入花期后,于晴天08:00—10:00取长度0.5~1.0 cm的幼小花蕾,参考张红梅等的方法[15]进行预处理、固定、水浴和染色等操作,切开子房取中心部位少量分生组织常规压片,观察有丝分裂中期分裂相进行染色体计数。
1.2.2 三倍体与父母本农艺性状和解剖学比较 观察记录F1三倍体与父母本植株的株高、冠幅、萼片等农艺性状并进行比较分析;将叶片背面表皮用镊子撕下在40倍显微镜下用带有标尺的目镜测量气孔大小[16];取同源三倍体矮牵牛F1植株成熟花粉粒于载玻片上,用1%醋酸洋红染色3~5 min,在光学显微镜下观察。以染色深、大小正常、外形圆的为可育花粉,不染色或染色浅、外形皱缩畸形的为败育花粉[17],统计可育花粉率,可育花粉率=可染花粉数/花粉总数×100%。
1.2.3 同源三倍体减数分裂观察 矮牵牛F1三倍体植株进入开花期后,于晴天08:00—10:00取长度4~7 mm的幼蕾,按照同源四倍体花粉母细胞减数分裂观察方法[18]进行。
2 结果与分析
2.1 同源三倍体的获得与子代染色体分布
获得同源三倍体杂种的途径通常都以同源二倍体、四倍体为亲本进行正反交。由表1可知,以矮牵牛四倍体07-12、二倍体梦幻为杂交亲本的正交坐果率为72%,反交坐果率为64%,授粉后结果率均较高,表明二倍体、四倍体正反交均能较好地完成授粉受精,子房能够正常生长膨大;而正反交果实中含有发育正常的种子数均很少,正交单果种子数1.2个,反交为1.0个,与四倍体07-12自交单果种子数73个、二倍体梦幻自交207个相差很大,这表明正反交胚胎在发育过程中绝大多数都发生了败育,不能够形成正常种子。
对杂交F1后代植株的染色体数目进行观察,由表2可知,正交后代中检测的9株中有6株为2n=21的三倍体(图1-1),还有2n=20、2n=22的非整倍体各1株和2n=28的四倍体1株;反交后代检测的6株中有3株为2n=21的三倍体,还有2n=22的非整倍体1株、2n=14的二倍体和2n=28的四倍体各1株。杂交后代中都出现了少量四倍体,正交是因为二倍体父本产生了少量2n花粉,反交中则为二倍体母本产生了少量2n卵细胞。
2.2 三倍体与父母本农艺性状和解剖学比较
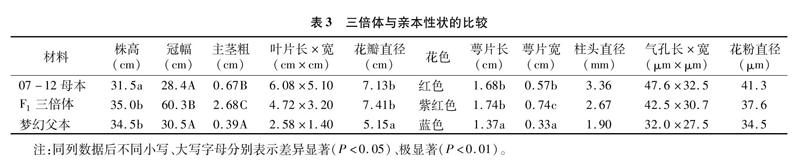
由表3可知,同源三倍体F1冠幅、主茎粗、萼片宽均显著或极显著大于双亲,而株高、花瓣直径、萼片长度偏向于数值较大的双亲中的一方,且与之差异不显著,叶片长度与宽度、柱头直径、气孔长度与宽度和花粉直径介于双亲之间,花色则与父母本都不相同(图2)。可育花粉率以F1三倍体最低,为44.7%,花粉外观形态有圆形、三角形和四边形(图1-2),二倍体梦幻最高,为95.2%,外观形态以圆形和三角形为主(图1-3),四倍体07-12为76.5%,以四边形为主(图1-4)。
2.3 三倍体花粉母细胞减数分裂观察
三倍体矮牵牛花粉母细胞的减数分裂过程基本符合二倍体植物减数分裂的规律,包括2次连续的核分裂和1次同时细胞质分裂,但也有一些异常现象。
前期Ⅰ的细线期染色体成细丝状(图1-5),双线期染色体开始缩短变粗,配对的同源染色体开始排斥分离,7个长线状染色体呈现0、U、N、S、9、ǒ等形状(图1-6)。终变期染色体进一步短缩,36.7%的细胞核内分布着7个同源配对的三价体, 多呈现棒状、U形或Y形(图1-7),其余大部分细胞核内都出现单价体、二价体和三价体混合共存的现象(图1-8),经统计平均单价体为3.8,二价体为3.2,三价体约为3.6。
中期Ⅰ:大多数细胞染色体排列在赤道板上(图1-9、图1-10),也有约6.2%的少数细胞出现游离在赤道板外染色体(图1-11)。通常在此期,二价体即将解离成2个单价体(图1-9),三价体将解离成二价体和单价体(图1-10)。
后期Ⅰ:二价体、三价体配对的同源染色体分别呈现 1 ∶1 和1 ∶2分离,由纺锤丝随机牵引至两极,未配对的单价体则随机牵引至两极中的任一极,因此导致两极的染色体数目不等。观察了318个花粉母细胞,有10/11(图1-12)、9/12(图1-13)、8/13(图1-14)和7/14(图1-15)多种分离方式,经统计,其比率分别为57.9%、32.4%、8.4%和1.3%。
后期Ⅰ中染色体不均等分离将导致各种非整倍体配子形成,为创制系列非整倍体提供了可能[19],其中可利用8/13分离通过回交直接获得初级三体[17],而7/14则是获得2n配子的细胞學基础[20]。后期Ⅰ姊妹染色单体一般不分离,但少数细胞出现姊妹染色单体提前发生分离(图1-14至图1-18),其中有位于同极的1对姊妹染色单体提前发生分离(图1-14),1对(图1-16)和2对(图1-17)姊妹染色单体提前发生分离分别牵引至两极,还有提前分离的姊妹染色单体(图1-18)和单价体(图1-19)滞留在主核外成为丢失染色体,丢失染色体将导致微核形成。
末期Ⅰ:存在落后和丢失染色体(图1-20)和染色体桥现象(图1-21)。
中期Ⅱ:大部分细胞2组染色体分别排列在各自的赤道板上,约有10.4%的细胞出现赤道板外染色体(图1-22)。
后期Ⅱ:大多数细胞中2组染色体的姊妹染色单体正常分离移向四极,观察到主要有10/10/11/11(图1-23)、9/9/12/12(图1-24)、8/8/13/13(图1-25)染色体分离方式,经统计,其比率分别为53.6%、37.1%、9.3%。
末期Ⅱ:存在落后和丢失染色体(图1-26)。
四分体时期:正常四分体比率约为86.7%,主要有四面体型和平行型2种类型(图1-27)。减数分裂过程中滞留在细胞质的丢失染色体往往导致微核形成,观察到具有1~2个微核的四分体(图1-28)。
对三倍体花粉母细胞减数分裂的中期Ⅰ、后期Ⅰ、末期Ⅰ、中期Ⅱ、末期Ⅱ和四分体6个时期染色体异常行为进行统计分析,结果表明,各时期平均约有8.6%的花粉母细胞减数分裂表现异常(表4)。
3 讨论与结论
获得植物同源三倍体的途径主要是通过二倍体、四倍体进行杂交,而二倍体、四倍体杂交不稔或稔性低是普遍存在的现象,如大白菜[21]、白菜[22]就未能获得同源三倍体, 谷子三倍体结籽率为0.24%[23]、甘蓝为3.61%[24]、菜薹仅为0.37粒/荚[25]。本试验中以二倍体、四倍体矮牵牛作亲本进行正反杂交均能够获得较高的坐果率,但单果结籽数均较低,表明正反杂交柱头能够接受花粉受精,子房能正常膨大,但胚胎发育成种子过程中均存在严重障碍。对菜薹胚胎学研究表明,三倍体胚胎败育是由于胚乳的衰退失去了养分来源,同时内珠被细胞迅速坏死阻塞营养物质的运输通道[25],矮牵牛三倍体败育具体原因还有待对胚胎的发育过程进行研究。
同源三倍体矮牵牛的花药大都能正常开裂,花粉较多,可育率为44.7%,作为父本与二倍体杂交都有较高的结籽率。一般认为,三倍体的育性与染色体基数有关,染色体基数少的物种产生具完整染色体组配子的概率较大,育性较高。但黄瓜和矮牵牛染色体基数都为x=7,而黄瓜同源三倍体花粉可育率仅有18.8%[17],谷子染色体组基数为x=9,甘蓝为 x=10,谷子三倍体几乎没有生活力花粉[23],甘蓝同源三倍体花粉生活力却高达65.49%[24],可见染色体基数数量不是决定因素。三倍体矮牵牛后期Ⅱ由染色体分离方式7/7/14/14形成完整染色体组花粉理论比率为(1/2)6=1.56%,而花粉可育率却为44.7%,表明许多非整倍性染色体数目的花粉具有生活力,可见植物三倍体的育性主要由对添加额外染色体的忍受程度决定,矮牵牛具体添加了哪些染色体的非整倍体花粉仍具有生活力尚有待进一步研究。
本研究获得的同源三倍体无论是自交还是与二倍体回交都有较高的结籽率,下一步将对杂交后代进行染色体计数和鉴定,从中筛选各种三体、单体等非整倍体,矮牵牛同源三倍体的成功获得为后续细胞遗传研究奠定了基础。
参考文献:
[1]李 健,王锦秀,王立英,等. 无籽枸杞新品种选育研究[J]. 西北植物学报,2001,21(3):446-450.
[2]张成合,王东平,申书兴,等. 菜薹部分初级三体的选育与细胞学鉴定[J]. 中国农业科学,2003,36(6):681-684.
[3]Ishiki K.Cytogenetical studies on African rice,Oryza glaberrima Steud.3.Primary trisomic produced by pollinating autotriploid with diploid[J]. Euphytica,1991,55(1):7-13.
[4]Khush G S,Singh R J,Sur S C,et al. Primary trisomic of rice:origin,morphology,cytology,and use in linkage mapping[J]. Genetics,1984,107(1):141-163.
[5]陈方永,倪海枝,徐小菊,等. 种间杂交培育“温岭高橙”三倍体杂种[J]. 浙江大学学报(农业与生命科学版),2013,39(5):531-536.
[6]刘文革,王 鸣. 西瓜甜瓜育种中的染色体倍性操作及倍性鉴定[J]. 果树学报,2002,19(2):132-135.
[7]赵艳华,程和禾,吴雅琴,等. 三倍体葡萄种质创新及倍性快速鉴定[J]. 园艺学报,2011,38(6):1161-1166.
[8]满 红,张成合,柳霖坡,等. 结球甘蓝二、四倍体间杂交三倍体的获得及细胞学鉴定[J]. 植物遗传资源学报,2005,6(4):405-408.
[9]吴志祥,聂京涛,曹 娴,等. 利用ISSR对矮牵牛品种遗传多样性的聚类分析[J]. 上海交通大学学报(农业科学版),2012,30(3):36-44.
[10]代色平,包满珠. 矮牵牛花药培养及植株再生研究[J]. 亚热带植物科学,2003,32(2):55-57.
[11]郭余龙,闫明旭,陈 君,等. 矮牵牛PMADS9基因启动子的克隆及分析[J]. 植物遗传资源学报,2011,12(2):275-280.
[12]杨 莎,张 彬,韩 垚,等. 矮牵牛PDS基因的克隆及其在shRNA介导的基因沉默中的应用[J]. 园艺学报,2017,44(2):315-322.
[13]李 颖,刘姬艳,胡江琴,等. 矮牵牛转基因体系的建立及转BIO和bio基因研究初报[J]. 浙江大学学报(农业与生命科学版),2013,39(1):42-49.
[14]魏 跃,王开冻,李洪海,等. 矮牵牛四倍体的诱导及其形态特征[J]. 江苏农业科学,2007(3):125-127.
[15]张红梅,张蜀宁,孔艳娥,等. 利用青花菜幼嫩雌蕊进行染色体核型分析[J]. 园艺学报,2009,36(5):727-730.
[16]张凌媛,郭启高,李晓林,等. 枇杷气孔保卫细胞叶绿体数目与倍性相关性研究[J]. 果树学报,2005,22(3):229-233.
[17]刁卫平,崔 利,江 彪,等. 黄瓜同源三倍体创制及减数分裂行为观察[J]. 西北植物学报,2009,29(1):36-42.
[18]魏 跃,张蜀宁,隋益虎,等. 同源四倍体矮牵牛花粉母细胞减数分裂观察[J]. 西北植物学报,2008,28(9):1742-1745.
[19]申书兴,侯喜林,张成合. 利用小孢子培养创建大白菜初级三体的研究[J]. 园艺学报,2006,33(6):1209-1214.
[20]钟 程,张蜀宁,于旭红,等. 白菜2n配子诱导及有性多倍化创制四倍体的研究[J]. 园艺学报,2010,37(11):1789-1795.
[21]刘学岷,申书兴,孙日飞,等. 大白菜二倍体与四倍体杂交后代倍性及胚胎学观察[J]. 园艺学报,1996,23(3):309-311.
[22]胡金良,徐汉卿,刘惠吉,等. 二倍体和四倍体小白菜的胚胎学研究[J]. 南京农业大学学报,1996,19(4):15-19.
[23]王润奇,高俊华,王志兴,等. 谷子三体系列的建立[J]. 植物学报,1994,36(9):690-693.
[24]柳霖坡,张成合,张 丽,等. 甘蓝异倍体间杂交获得三倍体和非整倍体的研究[J]. 河北农业大学学报,2003,26(3):69-72.
[25]張成合,张书玲,申书兴,等. ‘青露菜薹三倍体的获得及其胚胎学观察[J]. 园艺学报,2001,28(4):317-322.李刚波,陈 刚,赵 林,等. 大棚栽培对砂梨果实生长发育及品质的影响[J]. 江苏农业科学,2020,48(2):136-139.
