植物功能多样性及其在苔藓植物中应用的研究
2020-04-09张永昌郭水良
张永昌,何 林,郭水良*
(1.上海师范大学生命科学学院,上海200234;2.遵义师范学院生物与农业科技学院,贵州遵义563006)
苔藓植物是一类由水生向陆生过渡的、特殊的高等植物类群.由于苔藓植物对环境特别敏感,对水分和矿质营养有特殊的吸收方式,能够利用孢子进行远距离传播,并且有多种营养繁殖方式,很多苔藓植物耐旱能力强,使其能广泛分布于海洋以外的地球绝大部分生境中[1-5],是在种数上仅次于被子植物的高等植物类群.全世界苔藓植物约有23 000种[5],中国有150科591属3 021种,其中,藓类有86科431属1 945种,苔类有60科152属1 050种,角苔类有4科8属26种[6],是世界上苔藓植物多样性程度最高的国家之一.
苔藓植物具有众多的生物多样性价值,是温带和亚高山生态系统的主要组成成分,在水分涵养、物质循环、气候调节等方面发挥着重要作用,同时在环境监测、园林景观、药物开发等方面也至关重要.苔藓植物在生态系统的生态功能以及功能多样性,已经成为苔藓植物生态学研究的热点之一.
功能多样性最初定义为影响群落中生态系统功能的所有物种及有机物的功能性状值及其变动范围,强调的是性状的差异性[7].因其与生态系统功能关系紧密,功能多样性研究日益受到人们的重视[8].目前,国内外对植物功能多样性的研究主要集中在维管植物,已经比较成熟,研究领域包括:植物功能性状沿环境梯度的变化规律(与气候、海拔、地形等的关系)[9-16];植物功能性状对干扰(包括土地利用变化、火烧、放牧等)的响应[17-18];植物功能多样性与物种多样性或生态系统功能之间的关系[19-22].
PARDOW 等[23]基于苔藓植物生活型研究了法属圭亚那热带雨林树附生苔藓植物的功能多样性,以探究该地区苔藓植物多样性的热点区域.BERDUGO 等[24]在美国Whiteface 山的云杉林带研究了林窗处苔藓植物物种多样性和功能多样性,并创新性地使用苔藓植物的形态、生态和生殖的多种性状组合,研究了沿自然冠层开放梯度的苔藓植物功能差异.功能多样性已继物种多样性和系统发育多样性之后逐步成为研究苔藓植物多样性的又一重要指标,对开展苔藓植物多样性的研究和保护具有重要的意义.
基于此,本文作者对功能多样性在植物类群的研究进展,及苔藓植物功能多样性的研究成果进行了综述,并展望了未来苔藓植物功能多样性研究的方向.
1 植物功能多样性概念
相对于物种多样性,功能多样性更能反映植物对环境驱动力的适应以及植物内不同功能之间的权衡关系,也更能体现植物在生态系统中的作用.目前,关于植物功能多样性的研究涉及从低等植物到高等植物,从水生植物到陆生植物,从个体到群落等多个层次[14,25-28],但植物功能多样性的概念至今还没有标准的定义[29-31].TILMAN[7]将功能多样性定义为能够影响生态系统运作或行使功能的生物多样性的成分,但很难据此含义对功能多样性进行定量化的描述[30].目前普遍为人们所接受的功能多样性定义是指某一群落内物种间功能性状的变化范围或指特定生态系统中所有物种功能性状的数值和范围[31].
2 功能性状的选择
植物功能性状指影响植物生长、繁殖和生存的一系列植物属性[32],在种子植物中包括了植株高度、生活型、种子干重、比叶面积、叶形、叶片氮含量等,人们常根据研究问题的需要和植物类群选择不同的功能性状,如在讨论水分、温度对植物的影响时,会选择与蒸腾作用相关的比叶面积.在维管植物中,研究者常在一个或多个生态系统中,关注环境梯度与植物功能性状变化间的关系,探讨植物对环境变化的响应[15,33-34].与植物建立、生长、繁殖、传播和维持有关的性状也受到重视,这些性状包括叶片(比叶面积、叶片形状、叶片氮含量、叶片磷含量等)、植株(生长形态、株高等)、繁殖器官(花期长度、果实类型和成熟期、种子干重等),探讨这些性状在不同生境中的变异规律[11,14,16,35-38].苔藓植物生活史特殊,配子体发达,没有维管组织,又是变水植物,对环境特别是小生境变化非常敏感,在营养吸收和水分传导方面有自己特殊的方式,从而表现出与其他高等植物明显不同的生态适应特征.因此,在苔藓植物中开展功能多样性研究,人们常选择与水分、营养和光合生理关系密切的生活型,与环境水分利用有关的叶片细胞特征(疣、中肋、茎附属物等),和繁殖有关的孢子体特征(蒴齿长短、孢子大小等)[23,39-41].
3 功能多样性指标及其计算方法
目前还没有被普遍接受的功能多样性测度方法,研究者常根据研究的实际情况选择某种功能多样性指数来反映植物功能性状的变化[42-43].
MASON 等[44]依据物种多样性的组成,把功能多样性划分为功能丰富度(functional richness)、功能均匀度(functional evenness)和功能离散度(functional divergence).SCHLEUTER 等[31]在前人的基础上,对植物能多样性指数做了进一步的研究,通过对不同功能多样性指数的比较和分析,阐明了不同功能多样性指数的特征及其与其他指数的相关性及缺点,并基于此总结发展了新的功能多样性指数的计算方法.MCPHERSON 等[42]用R 语言编制了功能冗余指数的计算程序,并认为前人提出的功能多样性指标不能很好地反映性状数量、性状值性质、性状分布或丰度频率敏感性等,对这些敏感性的更好理解能使研究者将多样性指数与生态群落的基本特性联系起来,从而有助于选择最适合某一特定研究目的的指数.宋彦涛等[45]和陈又清[8]已对植物功能多样性及其运算方法做了相关的综述,在这里本文作者仅对常用的功能多样性指数及运算方法进行简要的介绍.
3.1 功能丰富度
功能丰富度是指物种或个体在群落中所占空间或功能范围的大小[42],是体现生态空间利用程度的指数.该指数越大,生态空间利用程度越高[46],用来计算某一群落该性状生态位空间占全部群落中该性状生态位空间的比例[44]:

其中,FRci为在群落i 中性状c 的功能丰富度;SFci为群落中物种所占据的生态位空间;Rc为所有群落中性状c占据的生态位空间.
3.2 功能均匀度
功能均匀度指数(FE0)用来表征物种性状在所占据性状空间的分布规律,该指数越大,说明物种分布得越规律,对有效资源的利用越全面[44].

其中,S 为物种数;PEWi'i+1为物种i和i+1 性状值权重差异的百分比;Ai为物种i的相对多度;Ci为物种i的性状值.
3.3 功能离散度
功能离散度指数(FDS)表示群落内有机体性状值的差异性,体现了群落内有机体的生态位分化和资源竞争程度,该指数越高,说明群落内有机体生态位互补程度越强,竞争越弱[44].
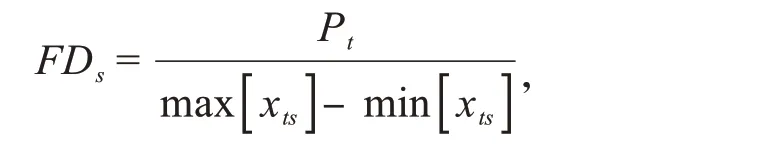
其中,Pt=Q1(Yt)-Q3(Yt);Yt为物种s 的性状平均值重复次数,Yt=[(Xts,Xts,…,Xts)×As];Xts为群落中每个物种的性状平均值;As为群落中物种s的多度;Q1和Q3分别为Yt的下四分位数和上四分位数.
在进行功能多样性指数计算时,推荐使用R语言中的“FD”Package和FDiversity软件进行计算.
4 维管植物功能多样性研究现状
理论上,植物周围的种种环境因子都会在不同程度上影响其生长发育.在功能多样性研究中比较关注气候(温度、水分、光照等)、土壤(pH 值、含氮量、含磷量等)、地形地貌(海拔、坡度、坡向等)、干扰(放牧、土地利用等)等因素.
(1)水作为植物生长的必需条件,对植物功能多样性有重要的影响.ALBRECHT 等[16]在Kilimanjaro山宽广的气候和土地利用梯度上,分析了200种植物花和果实性状的变化规律,发现研究区域植物的功能多样性主要受降水的限制.SAIZ等[15]以南极洲以外其他大陆的100种旱地植物为研究对象,通过对植物空间分布、株高和叶面积指数在环境梯度上的变化式样分析,发现降水量的季节性变化是影响群落功能多样性的主要因素.NUNES 等[11]以地中海旱地生态系统的橡树林地为研究对象,分析了13 个与植物建立、生长、繁殖、传播和种群维持有关的性状,发现功能离散度随着干旱程度增加而降低,从而增加了干旱区种间的功能相似性.ORDONEZ 等[35]以在广泛分布于欧洲的6 个被子植物类群为对象,估测了植物功能丰富度和离散度的地理格局,比较了叶面积指数、最大茎高、种子干重、茎比密度、生长型等5个功能性状,根据将第四纪冰川期—间冰期气候变化和当代环境因素(气候、生产力和地形)作为评价功能多样性决定因素的相对重要性,发现气候变化可能会导致植物功能多样性的长期失衡.
(2)土壤状况也与植物的功能多样性密切相关.ZHU 等[9]以中国黄土高原的16 个半干旱恢复草地为区域,研究了植被群落功能组成对土壤侵蚀的影响,定量计测了对恢复草地群落内13 个优势植物中的9 个可能与土壤侵蚀相关的功能性状,发现功能离散度与土壤侵蚀程度之间存在着较强的负相关关系,表明植物功能性状多样性的提高会减少该地区土壤侵蚀.ZUO 等[36]以半干旱草地的植物株高、比叶面积、比叶重、叶干重等性状值为依据,研究放牧和土壤性质诱导植物功能多样性对地上生物量和地下生物量的影响,发现放牧和土壤性质引起的生物多样性变化对草地生态系统功能的影响强烈依赖于植物功能多样性.张雪妮等[12]研究了土壤水盐变化对艾比湖荒漠植物群落水平的功能性状的影响,涉及植物形态、生理和化学等8 个功能性状,并从多维性状和一维性状角度进行分析,发现不同生活型植物间的功能性状在高水盐环境下差异显著,而在低水盐环境下趋于相似,揭示了功能多样性对土壤水分和盐分变化的响应规律.ZHANG 等[47]在研究废弃农田内植物的功能群组成、物种多样性与土壤性状的关系时,也得出了相似的结论.
(3)经纬度、海拔、坡向等地理因素通过影响水分、温度等间接地影响植物功能多样性,这种影响有明显的尺度效应[10].STEINERT 等[48]研究了挪威东南部虫煤植物群落的物种多样性和功能多样性,基于植物9 个花部特征计算了功能多样性指数,发现海拔会影响物种丰富度和功能均匀度.LIU 等[13]比较了29 个亚热带湖泊水生植物的物种丰富度和功能多样性,计测了物种丰富度和代表水生植物营养、再生和生态特征的13个功能性状,发现与低海拔区域湖泊相比,高原湖泊的功能丰富度较低,但功能均匀度和分散度较高,功能丰富度与环境变量之间无显著相关性,表明功能多样性对环境变量的抗性较强.ZHANG 等[10]研究了高原高寒草甸植物功能群组成、物种多样性与土壤性质(pH值、土壤总氮、土壤含水量)的关系,结果表明,植物群落特征和土壤理化性质在不同坡度水平上存在显著差异,坡面通过影响水分等生态因子和土壤理化特性,直接或间接地影响高寒草甸植物的物种组成和功能群差异.朱云云等[49]以黄土高原不同坡向上的自然草地群落为对象,选择了能很好反映植物竞争强度、响应环境变化的两个功能性状指标——株高和比叶面积,比较研究了植物功能性状和功能多样性指数随坡向的变化规律,通过比较黄土高原马栏地区山地的各坡向植物功能性状和3 种功能多样性测度指标的变化,发现阴坡和半阴坡的植物性状占据了更多的生态位空间,资源利用较充分且竞争较低,并且出现了高度的生态位分化;而阳坡和半阳坡由于水分等条件的限制,植物性状可占据的生态位空间有限,且在占据的生态位空间资源使用不足,导致生态位分化程度较低,资源竞争比较强烈,即阴坡对草地群落功能多样性的影响更高.
(4)干扰也会强烈地影响植物的功能多样性.研究者们先后研究了放牧、刈割、施肥等与植物物种多样性和功能多样性的关系[14,17-18,50].FORD 等[14]以英国威尔士西北部的两个原始自然林地和两个放牧林地为对象,用植物功能性状变化探索放牧强度和植物功能多样性间的联系,将比叶面积、叶片干重(LDMC)与植物耐受性指数(遮荫性、耐旱性和耐涝性)相结合,计算了群落加权平均值(CWM)和功能多样性;结合LDMC、平均株高、生长形态、光照、水分指标,计算了林下植物群落的加权平均值和功能多样性指数,发现与放牧林地相比,未放牧的林地其幼苗的建立,幼树的补充,和树级的功能多样性更强,表明控制放牧强度有利于林地生态系统恢复.YANG 等[17]在海口市内选择了461 个样地,基于花粉、种子、叶的相关性状,研究了土壤利用类型对植物功能多样性的影响,发现土地利用遗留效应对木本植物在分类和功能多样性上存在显著差异,其中以居住区的土地利用遗留效应最为明显,建议在快速城市化地区的保护项目中更多地关注居住区的植被,特别是低实用价值的物种.石明明等[18]以青藏高原高寒草甸中的17种共有植物为对象,利用比叶面积、株高、叶干物质含量、叶分配4种植物功能性状,分析了围封与放牧处理对植物功能性状和功能多样性的影响,发现不同植物功能性状对围封与放牧处理响应存在差异,放牧降低了比叶面积和植物高度,而叶干物质含量和叶分配在处理间无显著差别,反映了植物对资源获取和利用存在权衡.放牧在总体上降低了植物功能性状(比叶面积、叶干物质含量、叶分配、植株高度)的种间平均差异及分离度,反映了植物对光资源的竞争减弱.该结果与孔彬彬等[50]、陈超等[51]等关于刈割对高寒草甸物种多样性和功能多样性关系的影响的实验结果一致,放牧可能导致植物对地上资源的竞争转变为对地下资源的竞争.因此,在高寒草甸地区,今后关于植物功能性状多样性的研究应加强地下过程相关性状的研究.WANG 等[52]、DICKIE 等[53]、ZUIDAM 等[54]分别阐述了入侵种或引进外来种对本地植物功能多样性的影响.UYTTENBROECK 等[55]基于生态位,发现食草动物取食行为会显著地影响生态系统的植物功能多样性.
5 苔藓植物功能多样性研究现状
功能性状的选择是功能多样性研究的基础.已有部分学者开展了苔藓植物功能性状的研究.例如,叶吉等[56]通过对长白山北坡暗针叶林地面和树附生苔藓植物生物量的研究,发现苔藓植物生物量及物种组成能够很好地指示长白山北坡林型,生物量能够很好地指示苔藓植物的生态功能;杨武等[57]对源于不同生境的17 种藓类植物叶片进行了比较解剖,发现藓类植物中肋导水主细胞的有无、厚壁细胞是否分化、中肋细胞层数和密度、叶片细胞层数和密度、叶表附属物等差异显著.WANG等[58]比较了川西老龄冷杉林下28 种苔藓植物的生长形态以及与光合作用相关的生理指标,发现12 种直立型和16 种平伏型苔藓单位质量的氮与最大光合能力回归关系的斜率相同,但前者将更多的氮用于合成光合色素,而后者的形态结构更有利于捕获光能;同时发现苔藓植物生长形态对其营养分配和光合效率有很强影响.SIERRA 等[59]以苔藓植物生殖性状为指标,预测叶附生苔藓植物群落的形成时间.WANG 等[60]研究了西藏干旱/半干旱地区苔藓植物形态特征及其对环境变化的响应,发现苔藓植物的形态特征与环境变化密切相关,不同基质环境变化对苔藓植物的影响差异较大,苔藓植物的各种功能性状密切相关,这些性状的组合对环境变化有协同反应.
近年来,苔藓植物群落的功能多样性也开始受到关注,应用的功能特征有配子体形态[39]、生活型[23],以及与水分吸收保持有关的性状[40]等.AH-PENG 等[39]应用苔藓植物形态特征,发现La Réunion 岛亚高山苔藓植物的功能多样性与物种多样性存在正线性相关,地表分布的苔藓植物群落具有更高的功能冗余,比树附生苔藓植物群落具有更强的抗干扰能力.基于与水分获取和保持有关的功能性状,HENRIQUES 等[40]研究了Azorean 苔藓植物功能多样性和种类构成特点,发现沿海和山地苔藓植物在功能性状构成上明显不同,气候变化不仅会影响苔藓植物种类的组成,也会影响苔藓植物的功能多样性.PARDOW 等[23]基于生活型研究了法属圭亚那热带雨林树附生苔藓植物的功能多样性,发现低地雨林是该研究地区的苔藓植物多样性热点区域.LISBOA等[41]研究发现随着海拔的上升,藓类植物孢子变大,叶片变长,高度增加,功能多样性提高.BERDUGO 等[24]在美国Whiteface山的云杉林带,创新性地尝试使用苔藓植物的形态、生态,以及生殖的多种性状组合,研究了沿自然冠层开放梯度的苔藓植物的功能差异,发现林窗大小与物种多样性之间不存在显著相关性,而指示种和植物功能性状可以用来评价沿环境梯度的苔藓植物群落生态的变化.
6 苔藓植物功能多样性研究展望
目前尚未检索到国内关于苔藓植物在不同生态系统中功能多样性研究工作的报道,这使得我国的苔藓植物多样性保护策略仅立足于物种数量状况,而没有全面考虑物种的功能性状,不能够全面地认识苔藓植物在生态系统中的作用和价值.因此,在国内开展苔藓植物功能多样性研究对于国内苔藓植物多样性的保护意义重大.
本文作者认为,在进行苔藓植物功能多样性研究时,可以根据苔藓植物的特点,将其功能特征分为以下4个方面:
(1)苔藓植物的生活型与水分、营养及光合生理关系密切,也影响着环境的湿度等,因此是重要的功能性状.常见苔藓的生活型主要有丛集型(Turf)(如金发藓、真藓等)、垫状(Cushion)(如紫萼藓等)、交织型(Weft)(如青藓、灰藓、羽藓等)、树状(Dendroids)(如树藓、万年藓等)、悬挂型(Pendant)等.另外,由于水分条件变异较大,有些适应能力较强、分布较广的种类(如木藓)在不同环境中发育生长成为不同的生活型;还需考虑植株密度、枝叶排列等指标.
(2)苔藓植物的繁殖特征,包括:有性繁殖(有无孢蒴)、无性繁殖(有无芽胞等营养繁殖结构)、有无蒴齿、蒴齿长短(与孢子传播有关)、孢子大小、蒴柄长短、苞叶大小、是否隐蒴等.
(3)样地中苔藓植物群落的盖度、高度是与生物量相关的指标,也具有重要的生态功能和生物多样性价值.
(4)苔藓植物的形态特征,主要包括:茎中央是否有导水细胞分化,是否有储水细胞,叶片先端是否有透明毛尖,叶片细胞是否具疣及发育程度,叶片细胞大小,叶片细胞是否多层,叶片干燥时是否强烈背卷,叶片干燥时是否强烈扭折卷缩,叶表是否有栉片,有无叶片中肋,茎上是否有附属物等,这些性状是否与环境水分利用、截留和保持有关.
在亚高山苔藓林、云冷杉林等寒温带森林生态系统,泥炭藓沼泽湿地等生态系统中,苔藓植物是重要的组成部分,今后应该选择具有代表性的生态系统开展苔藓植物功能多样性、物种多样性、系统发育多样性间关系的研究,还应该研究苔藓植物功能多样性对维持健康的生态系统的作用.
