基于多模型集合预测尖萼红山茶物种分布
2020-04-09陈思斯刘想童鑫玥管毕财
陈思斯, 刘想, 童鑫玥, 管毕财
基于多模型集合预测尖萼红山茶物种分布
陈思斯, 刘想, 童鑫玥, 管毕财*
南昌大学生命科学学院, 南昌 330031
气候是影响生物多样性和物种分布的一个重要因素, 近年来由于温室气体的大规模排放, 气候正在快速的发生变化, 对植物的分布影响巨大。研究采用山茶科山茶属尖萼红山茶(Hance.)作为研究对象, 通过实地考察和网上查阅获取其分布数据, 从世界气候网站和兰州寒区旱区科学数据中心获取环境变量, 结合R语言、Erdas和ArcGIS等软件对其当前和将来(2070)的分布区进行模拟并得出置信度和计算其面积变化和质心变化。研究结果表明, AUC值均大于0.9, 属于非常好的预测, 影响其分布的主要因子是Bio01(年平均温度)、Bio04(温度季节变化)、Bio15(降水量变异系数)和Bio03(昼夜温差与年温差比值)。在四种二氧化碳排放情景下(RCP2.6、RCP4.5、RCP6.0及RCP8.5)所进行的预测均表明尖萼红山茶的分布面积均有收缩, 且收缩面积随着碳排放程度的加剧而增加, 且最大收缩面积占总面积的80%。通过对该物种质心变化的预测中发现, 其总体呈现向北部迁移的趋势。通过本次的预测发现气候变化在未来可能对尖萼红山茶的生存威胁很大。
尖萼红山茶; biomod2; 物种分布模型; 面积变化; 质心变化
0 前言
气候和环境对生物的影响非常显著[1], 目前关于气候和环境对物种分布格局的定量研究已经成为了研究热点之一[2-3]。当前, 气候正快速的发生变化, Dawson 等人研究认为气候变化是威胁21世纪生物多样性的重要因素之一[4]。Coetzee等的研究表明极端天气事件频率增加, 会影响本土和入侵物种的分布[5], Wilson等的研究表明全球呈现总体变暖的趋势, 这将影响一些物种的北部栖息地范围[6]。近年来, 考虑气候变化对物种分布和生态位转移的潜在危害已成为生物多样性保护主义者的主要课题之一[7].
得益于近些年气候变化情景的深入研究和地理信息系统(GIS)的飞速发展, 我们可以利用物种分布模型来预测不同物种在气候变化下的适宜生境变化来解决景观生态学和保护生态学相关的热点问题[8]。物种分布模型(SDMs)是通过关联物种的分布记录生物气候变量研究物种与环境之间的关系, 并将这些关系投影到不同的空间和(或)时间预测目标物种适合的栖息地和潜在分布范围的模型[9], 目前该模型广泛应用于各种气候变化情景下不同物种的生境预测研究。主要包括MaxEnt、GRASP、GARP、Diva-GIS、Biomapper和SAM等模型, 但是使用单一的物种分布模型受限于模型算法的精确度, 物种和环境之间的不确定性等各种原因, 降低了单一模型的准确性[10], 所以多模型集合预测方法的使用就可以大大的提高预测精度, Biomod2[11]是R语言中的程序包, 其利用组合模型在构建物种分布模型中被广泛使用。雷军成等利用Biomod2通过模拟黑麂在气候变化情景下适宜生境的可能变化, 发现气候变化将导致黑麂适宜生境面积大幅度减少[12]。Sergio Noce等人通过使用Biomod2软件包研究南欧气候变化对南欧的森林物种的适宜性分布和多样性的变化可能性, 结果表明, 随着气候的变化, 大部分地区的物种多样性将减少, 而阿尔卑斯地区存在成为物种迁移避难所的潜力[13]。
尖萼红山茶(Camellia edithae Hance.), 属山茶科(Theaceae)山茶属(Camellia)灌木或者小乔木, 是中国的特有种, 该物种主要分布于广东省东部、江西省南部和福建省等南岭以南大部地区, 这些地区气候均温暖湿润, 但目前对尖萼红山茶的适应生境研究很少, 已发现的尖萼红山茶的分布点也极为稀少, 本研究希望通过运用组合模型的预测方式, 来初步探索尖萼红山茶可能的分布范围以及适宜其生存的环境情况, 并希望通过组合模型预测基于未来环境变化情景下尖萼红山茶的分布变化情况, 希望能为该物种的保护起到预警的作用。
1 材料与方法
1.1 物种分布数据
尖萼红山茶的主要分布信息来源于中国数字植物标本馆(CVH, http: //www.cvh.ac.cn/)、中国植物志(FRPS, http://www.frps.eflora.cn/)和野外调查。对于有详细地址和经纬度的标本记录, 我们使用Google earth进行校对, 采用准确无误的标本分布信息, 此外, 对于有详细位置但未记录经纬度的地点, 也使用Google earth进行定位并记录使用, 对于年代久远且地址不详细的分布点进行剔除, 此外, 为减少标本之间存在空间自相关, 使用ArcMap剔除部分距离过近和重复采样标本, 最终获得尖萼红山茶的分布数据共40个。图1即为尖萼红山茶的分布点示意图。
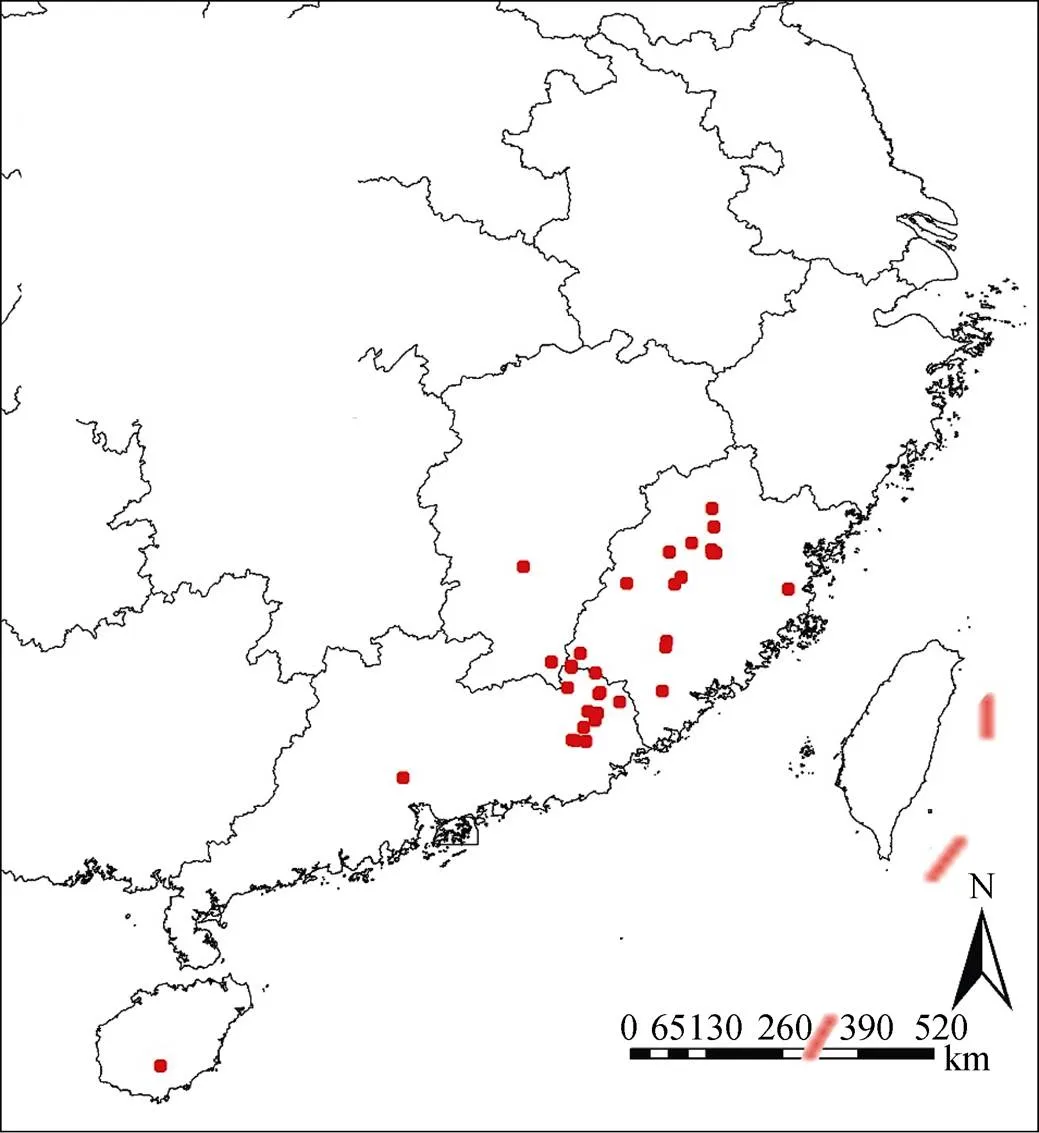
图1 尖萼红山茶分布点示意图
Figure 1 Distribution of Camellia edithae
1.2 环境数据
环境数据包含生物气候因子和数字高程模型数据。生物气候因子下载自世界气候数据库(www.worldclim.org), 选择当前(Current)和2070年两个时期的气候数据。气候数据来自于2014年第五次IPCC评估的气候模型。因为不同的气候模型的未来预测存在一些区别[14], 所以2070年的气候数据选择了对我国气候模拟时比较有优势的通用气候系统模型CCSM4[15], 每个气候模型选择RCP (Representative concentration pathways)2.6, RCP4.5, RCP6.0, RCP8.5四种二氧化碳排放情景。其中RCP2.6表示人类采用更加积极的方式应对气候变化, 使得温室气体排放下降并在本世纪末将其降为负值, 到本世纪中叶(2046—2065年), 全球平均地表气温上升0.4—1.6°C; RCP8.5表示假定能源消费结构改善比较缓慢, 温室气体保持较高速率的排放。在此情景下, 到本世纪中叶, 全球平均地表气温升高1.4—2.6°C。而RCP4.5和RCP6.0则介于二者之间, 到本世纪中叶, 全球平均地表气温分别升高0.9—2.0°C和0.8—1.8°C[16]。在所选的所有情景中, 为避免气候因子相关性较高导致共线性, 对其进行相关性分析(correlation coefficient), 排除相关性大于0.8的气候因子, 最终选择Bio01: 年平均温度、Bio02: 平均温差、Bio03: 昼夜平均温差/年温差、Bio04: 季节性温度、Bio13: 最湿月降水量、Bio14: 最干月降水量和Bio15: 季节性降水共7个生物气候因子参与建模。数字高程模型数据下载自兰州寒区旱区科学数据中心(http://westdc.westgis.ac.cn/)。所有环境数据均处理为2.5arc-min, 单个栅格面积大小约为22 km2, 分析底图采用1:400万中国行政区划图, 下载自国家基础地理信息系统网站(NGCC, http://ngcc.sbsm.gov.cn/), 地理坐标系为WGS1984。
1.3 模型的建立与评估
物种分布模型的建立需要物种的分布数据和缺失数据, 但由于并不能够获取到准确的物种的缺失数据, 故通常使用假缺失来代替物种真实缺失的数据[17-18]。我们采用Biomod2软件包进行不同模型的组合预测[19]。表2是Biomod2程序包中的十种不同算法。

表1 19个环境变量
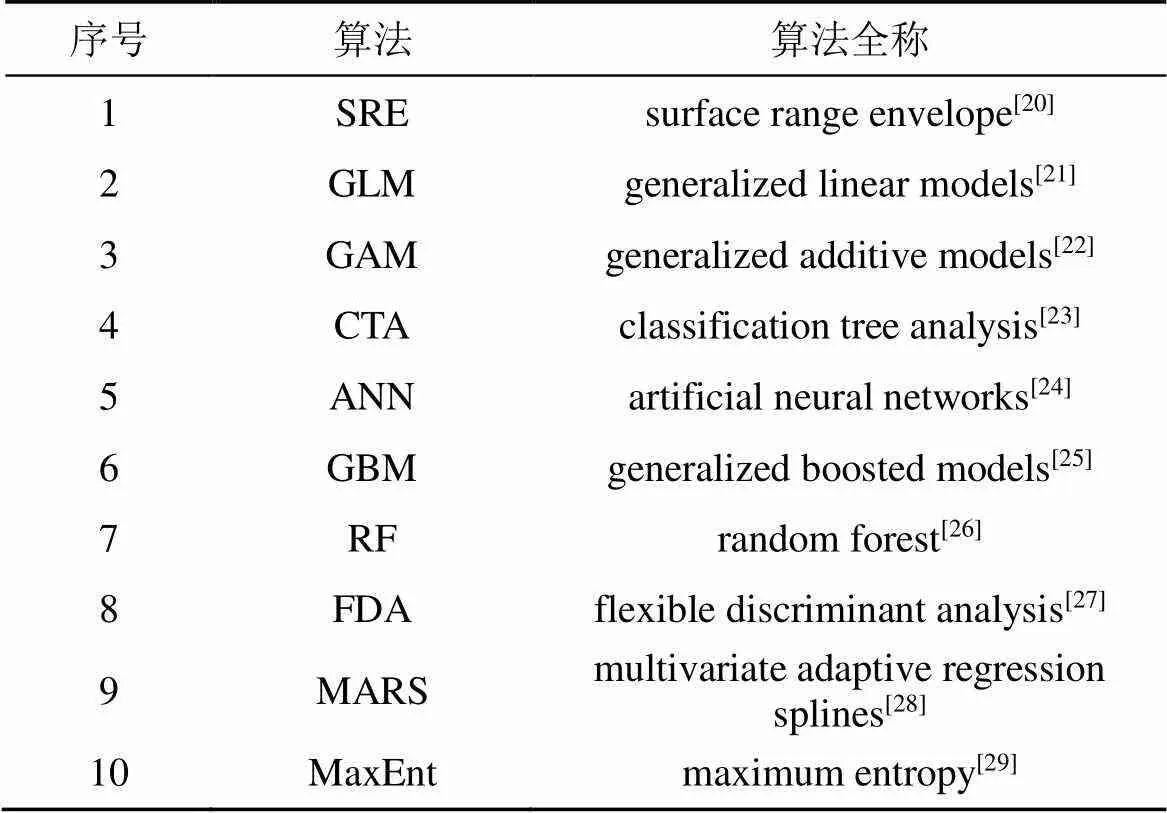
表2 参与模拟的10种算法
利用Biomod2中的十种算法, 使用已知物种分布数据的80%用于建模, 20%用于模型验证, 每个模型运行三次重复[30]。本研究选择Biomod2内嵌的两种模型评价指标来进行拟合精度的评价。即TSS和AUC。TSS是真实技巧统计法(True skill statistic), 综合了敏感度和特异度两个指标, 考虑了疏忽和替代性的错误。其取值范围在-1到1之间, 介于0.8到1之间表示拟合效果极好, 取值介于0.6到0.8之间表示拟合效果好, 取值介于0.4到0.6之间代表拟合效果一般[31-32]。AUC是指接收曲线ROC(Receiver operating characteristic)与横坐标围成的面积值, AUC值介于0.5—1之间, 当结果大于0.7时表示预测结果一般, 当结果大于0.8时表明结果满意, 当结果大于0.9时代表预测结果非常好。在物种分布模型的评价中, AUC得到了广泛的应用[33]。在运算过程中, 自动挑选TSS值大于0.7的算法进行构建组合模型运算, 最终使用AUC值作为预测结果的评价标准, 将预测结果的组合模型分别导入Erdas进行转换为Tiff格式, 使用Georeferencing工具对Erdas输出的不含坐标的文件进行地理配准。最后根据适宜度高低来进行可视化表达。
1.4 潜在分布范围及其变化的预测
将组合模型结果通过ArcMap转化为二进制SDM图层, 在转化过程中选择固定阈值法, 将大于0.5的栖息地划分为适宜生境, 反之则为不适宜生境。定义投影坐标系为WGS1984。在ArcMap中使用插件SDM tools中的Distribution change between binary SDMs工具进行不同时期生态位面积变化计算; 使用Centroid changes计算其质心变化, 并加入比例尺和指北针后导出。其中, 面积变化主要采用相减方式得出, 当前到未来由未来减去当前分布得出, 再通过ArcMap将结果可视化表达。
2 结果与分析
2.1 模型预测精度
通过对尖萼红山茶分布格局的模拟, 通过对单一模型与组合模型的AUC值可以看出(表3, 表4), 其单一模型的AUC值大都低于组合模型, 故得益于组合模型的使用, 不同时期的模型AUC值均高于0.998, 属于非常好的预测, 表明预测结果能够非常好的反映尖萼红山茶的当前及2070年的分布。不同时期AUC值见表3。
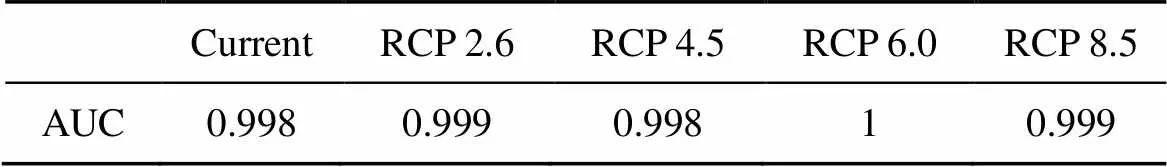
表3 不同时期的不同模型的AUC值

表4 不同时期的单一模型的AUC值
图2是十种算法在当前气候环境下的各个变量重要性, 从图中可以看出, 在八种环境图层中, 对尖萼红山茶物种分布模型影响比较大的是: Bio01 (年平均温度)、Bio04(温度季节变化)、Bio15(降水量变异系数)和Bio03(昼夜温差与年温差比值)。从图中可以发现结果的标准差均大于0.3, 造成这种结果的原因可能是因为不同的环境因子对不同的算法影响是不一致的[34]。
2.2 尖萼红山茶的分布格局模拟
通过ArcGIS对尖萼红山茶的栖息地适宜度进行可视化表达, 投影坐标系选择的是WGS1984, 图3为尖萼红山茶当前时期的栖息地适宜度, 颜色由蓝到红依次代表适宜度由低到高, 由图中可以看出最适合其生存的地区包括云南省东部, 广东和福建交接处。当前的气候模型条件下, 适宜分布区面积为436154km2(图3)。
2.3 尖萼红山茶的分布格局变化范围
本研究探讨了在未来(2070)的四种不同的温室气体排放情景下的尖萼红山茶分布格局的变化, 图4中A、B、C和D分别表示RCP2.6、RCP4.5、RCP6.0和RCP8.5的分布分局与当前分布格局之差, 即变化情况, 其中-1(紫色区域)代表扩张范围, 0(蓝色区域)代表现在和将来都不会分布的范围, 1(黄色区域)代表当前和将来都存在的范围, 2(红色区域)代表分布收缩的范围。
由图4中可以直观的看出, 尖萼红山茶在不同的温室气体排放情景下的分布范围均呈现收缩的趋势, 但是不同的情景区别较大, 具体表现为温室气体排放的越大, 则对其生存越不利。在RCP2.6情景下, 面积净收缩67738 km2; RCP4.5情景下, 面积净收缩123356 km2; RCP6.0情景下, 面积净收缩293133 km2; 而在RCP8.5情景下, 面积净收缩达到了350341 km2, 占到当前分布面积的80.33%。
2.4 尖萼红山茶质心变化
根据ArcGIS 10.2中SDM工具包中Centroid changes工具对不同温室气体排放情景下尖萼红山茶的分布质心变化进行分析, 底图利用中国地图进行导出, 结果见图4。
由图4中可以看出, 不同的温室气体排放情景下, 尖萼红山茶的质心变化大体趋同, 都向东北方向移动,但是随着碳排放的变化, 又各有不同, 具体表现为碳排放的程度越激烈, 则质心变化越大, 主要是从广东省向江西和湖南方向移动。而红色越深的区域表明核心分布范围内的高重叠区域随着时间的变化, 而推移。
3 讨论
3.1 影响尖萼红山茶分布的主要因子
本研究采用的环境变量为从世界气候网站下载的19个不同气候因子, 为了排除共线性影响后选取其中7个气候因子和数字高程模型, 模拟结果的环境因子相对贡献度显示, 海拔作为筛选后的八个环境因子之一, 是对尖萼红山茶的分布影响最小的一个因子。在对其分布影响排名中, 位居前四的是Bio01(年平均温度)、Bio04(温度季节变化)、Bio15(降水量变异系数)和Bio03(昼夜温差与年温差比值), 这四个环境因子均为温度和降水量, 结合其分布的环境可以发现是该物种分布在我国南部亚热带区域的原因, 这也符合我们之前所提到的温度和降水量对其影响最大的猜测。本研究基本与尖萼红山茶的实际分布吻合且符合气候因子对其生境影响的预期, 但是仅使用海拔和气候作为环境因子略有不足, 以后的研究中应该使用更多的环境因子参与计算, 例如有研究使用土壤作为影响因子之一参与模拟, 用以推测野生油茶()的潜在分布[35]。也有研究使用土壤pH作为影响因子参与模拟, 用以推测热带稀树草原树的分布受气候变化的影响[36]。还有研究使用坡度和太阳辐射等信息作为影响因子, 用以推测环境变化对狐猴科种群范围变化和保护的挑战[37]。因此结合这些文献我们选取了上述的8个环境因子来进行模拟, 这也是在以后的研究中要必须考虑到的。

图2 主要环境变量对预测尖萼红山茶分布的相对贡献率
Figure 2 Relative contributions of the main environmental variables to the model for

图3 当前时期尖萼红山茶潜在分布图(暖色颜色越深表示适宜程度越高, 红色区域表明适宜程度最高, 深蓝色区域表明适宜程度最低)
Figure 3 Camellia edithae potential distribution map in current (The warmer color means higher suitability, the red areas show the highest suitability and dark blue areas show the lowest)

图4 不同情景下尖萼红山茶的分布面积变化(-1表示适宜区域扩张范围, 0表示在和将来都不会分布的范围, 1表示当前和将来都存在的范围, 2表示分布收缩的范围)
Figure 4 Distribution change ofCamellia edithaein different scenarios (-1 means the expanding regions, 0 means no occupational place, 1 means the area with no change, 2 means the contractive place)
研究结果表明, 在云南省也有部分范围适宜尖萼红山茶的分布, 这与我们调查和收集到的实际分布点有些出入, 属于预测适宜生境范围大于已知分布范围, 造成这一现象可能有以下几点原因: 1、该地区确实有尖萼红山茶分布, 但是受限于调查力度和收集资料有限的缘故, 这些种群尚未被发现或者记录[38]; 2、地理壁垒导致尖萼红山茶并没有扩散到所有的适宜分布区。
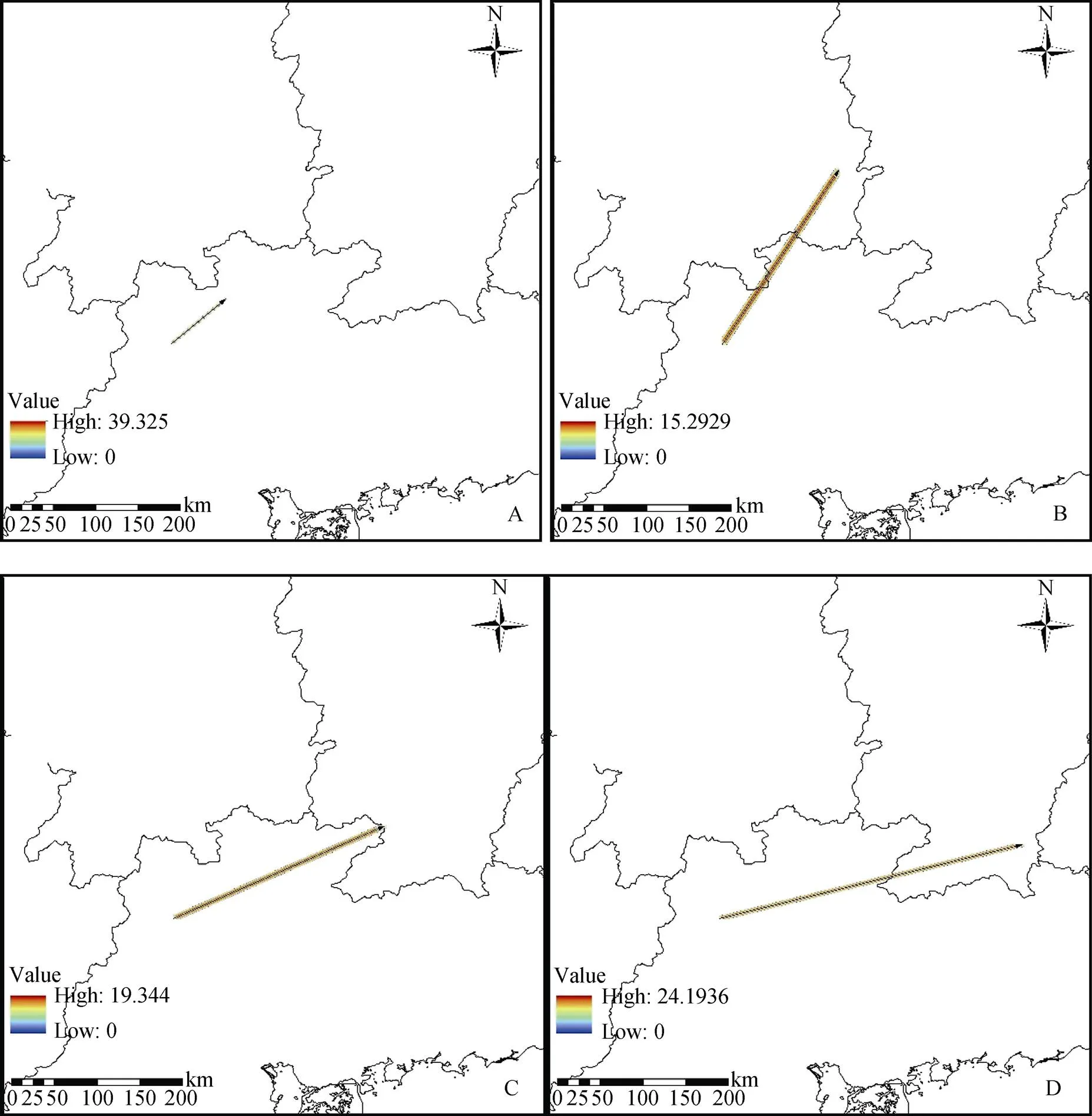
图5 不同情景下尖萼红山茶分布面积质心变化图 (图例表示线的密度, 较暖颜色表示的线密度说明了核心范围内的高重叠区域随着时间的推移而变化)
Figure 5 The variation of the centroid of the distribution area of Camellia edithae under different scenarios (The legend means line densities, the warmer colors illustrate areas of overlap in core range shifts through time)
3.2 尖萼红山茶的未来分布
结合环境因子对尖萼红山茶影响和未来的分布区域变化可以知道, 由于其受温度影响较大, 在温室气体排放的大背景下, 全球气温的是升高是必然趋势, 尖萼红山茶的适生区范围大幅度缩小也是必然趋势, 并且缩小范围会随着碳排放的增多而增大, 最保守的碳排放RCP2.6是其适生区最乐观的变化范围, 即缩小范围是67738 km2, 主要是南部区域适生区的减少, 同时北部也有少部分扩张; 在RCP4.5情景下, 原本适合生存的广东省大部分地区和云南省部分区域将不再适合其生存, 同时向北扩张, 在湖南省、江西省和浙江省部分地区得到生存的适宜条件。而在碳排放最剧烈的RCP8.5情景下, 超过百分之八十的分布区域将消失不见, 仅剩下武夷山部分区域尚适合其生存, 可能因为其典型亚热带季风气候而适合作为尖萼红山茶的避难所。因此, 合理控制碳排放是利于尖萼红山茶的生存与分布的。
随着气候变化, 基于全球气温升高的背景下, 尖萼红山茶的分布区域总体向北方迁移的趋势, 尖萼红山茶的质心变化则具体表现为向江西省和湖南省方向移动, 甚至是即将到达了福建省范围内。这和许多同类研究结果非常相似。有使用Biomod2建模并使用三种不同的气候模型研究气候变化对印度水稻的分布结果表明, 气候变化会导致水稻适生区范围向北部扩张[30]。有研究者使用MaxEnt建模探讨气候变化对中国八种入侵杂草的影响, 研究结果也表明, 我们需要采取适当的措施去控制杂草向北部扩张的趋势[39]。同时, 在全球范围内的一项关于气候变化对河蚬()的分布的研究中, 作者采用Biomod2建模探讨不同时期的气候变化对河蚬分布的影响结果也表明气候的变化将导致河蚬的生态位有向北移动的趋势[34]。尖萼红山茶与此类研究相似, 结果都表现出向北扩张的趋势, 但是我们认为这种趋势是由于南部适生区大范围收缩, 导致其分布质心往北移动的假象, 而非适生区往北扩张的结果。
总的来说, 得益于组合模型的应用, AUC值均大于0.95, 模拟精度较好; 对于尖萼红山茶而言, Bio01(年平均温度)、Bio04(温度季节变化)、Bio15(降水量变异系数)和Bio03(昼夜温差与年温差比值)这四个环境因子影响相对较大, 海拔因素对其影响则相对较小; 伴随着环境因子的变化, 尖萼红山茶的分布在未来的时间内将面临很大面积的收缩, 且总体向东北方向移动。
4. 结论
通过山茶科山茶属尖萼红山茶(Camellia edithae Hance.)当前和将来(2070)的潜在适宜分布区进行模拟并得出置信度和计算其面积变化和质心变化。研究结果表明, 可能影响其分布的主要因子是Bio01 (年平均温度)、Bio04(温度季节变化)、Bio15(降水量变异系数)和Bio03(昼夜温差与年温差比值)。在未来四种二氧化碳排放情景下(RCP2.6、RCP4.5、RCP6.0及RCP8.5)所进行的预测均表明尖萼红山茶的分布面积有收缩, 且收缩面积随着碳排放程度的加剧而增加。通过对该物种质心变化的预测中发现, 其总体呈现向北部迁移的趋势。气候变化在未来可能对尖萼红山茶的生存存在很大威胁。
[1] WILLIS K J, BHAGWAT S A. Biodiversity and Climate Change[J]. Science, 2009, 326(5954): 806–807.
[2] FITZPATRICK M, GOVE A, SANDERS N, DUNN R. Climate change, plant migration, and range collapse in a global biodiversity hotspot: the Banksia (Proteaceae) of Western Australia[J]. Global Change Biology, 2010, 14(6): 1337–1352.
[3] SAX D F. Saving a Million Species: Extinction Risk from Climate Change[J]. Ecological Restoration, 2001, 31(1): 99–100.
[4] DAWSON T P, JACKSON S T, HOUSE J I, et al. Beyond predictions: biodiversity conservation in a changing climate[J]. Science, 2011, 332(6025): 53–58.
[5] COETZEE B W T, KREMEN C, CAMERON A, et al. Conservation with Caveats[J]. Science, 2008, 321(5887): 340–342.
[6] ROBERT J W, DAVID G, JAVIER G, et al. Changes to the elevational limits and extent of species ranges associated with climate change[J]. Ecology Letters, 2005, 8(11): 1138–1146.
[7] GAMA M, CRESPO D, DOLBETH M, ANASTÁCIO P M. Ensemble forecasting ofworldwide distribution: Projections of the impact of climate change[J]. Aquatic Conservation: Marine and Freshwater Ecosystems, 2017, 27(3): 675–684
[8] WIENS J A, STRALBERG D, JONGSOMJIT D. Niches, models, and climate change: Assessing the assumptions and uncertainties[J]. Science, 2009, 106(17): 19729–19736.
[9] ELITH J, LEATHWICK J R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time[J]. Annual Review of Ecology, Evolution, and Systematics, 2009, 40(1):677–697.
[10] 张雷, 刘世荣, 孙鹏森, 等. 气候变化对物种分布影响模拟中的不确定性组分分割与制图——以油松为例[J]. 生态学报, 2011, 31(19): 5749–5761.
[11] WILFRIED T. BIOMOD - optimizing predictions of species distributions and projecting potential future shifts under global change[J]. Global Change Biology, 2003(10): 1353–1362.
[12] 雷军成, 王莎, 王军围, 等. 未来气候变化对我国特有濒危动物黑麂适宜生境的潜在影响[J]. 生物多样性, 2016(12): 1390–1399.
[13] NOCE S, COLLALTI A, SANTINI M. Likelihood of changes in forest species suitability, distribution, and diversity under future climate: The case of Southern Europe[J]. Ecology & Evolution, 2017, 7(22): 9358–9375.
[14] JAYASANKAR C B, SURENDRAN S, RAJENDRAN K. Robust signals of future projections of Indian summer monsoon rainfall by IPCC AR5 climate models: Role of seasonal cycle and interannual variability[J]. Geophysical Research Letters, 2015, 42(9): 3513–3520.
[15] 田芝平, 姜大膀. 不同分辨率CCSM4对东亚和中国气候模拟能力分析[J]. 大气科学, 2013, 37(1): 171–186.
[16] RIAHI K. Fifth Assessment Report[R]. IPCC, 2014.
[17] JIMÉNEZ-VALVERDE A, HORTAL J. Challenging Species Distribution Models: The Case ofin the Iberian Peninsula[J]. Annales Zoologici Fennici, 2008, 45(3): 200–210.
[18] LIU Xuan, ROHR J R, LI Yiming. Climate, vegetation, introduced hosts and trade shape a global wildlife pandemic [J]. Proceedings Biological Sciences, 2011, 280(1753): 20122506.
[19] THUILLER W, LAFOURCADE B, ENGLER R, et al. BIOMOD-a platform for ensemble forecasting of species distributions[J]. Ecography, 2010, 32(3): 36 9–373.
[20] BUSBY J R. BIOCLIM: a bioclimate analysis and prediction system[J]. Plant Protection Quarterly, 1991: 6.
[21] NELDER J A, WEDDERBURN R W M. Generalized Linear Models[J]. Journal of the Royal Statistical Society, 1972, 135(3): 370–384.
[22] HASTIE T, TIBSHIRANI R, BUJA A. Flexible Discriminant Analysis by Optimal Scoring[J]. Publications of the American Statistical Association, 1994, 89(428): 1255–1 270.
[23] LOH W. Classification and regression trees[J]. Wiley Interdisciplinary Reviews: Data Mining and Knowledge Discovery, 2011, 1(1): 14–23.
[24] BILLFULKERSON. Pattern Recognition and Neural Networks[J]. Technometrics, 1997, 39(2): 233–234.
[25] FRAYMAN Y, ROLFE B F, Webb G I. Solving Regression Problems Using Competitive Ensemble Models[J]. Springer Berlin Heidelberg, 2002: 511–522.
[26] BREIMAN L. Random Forests[J]. Machine Learning, 2001, 45(1): 5–32.
[27] HASTIE T, TIBSHIRANI R, BUJA A. Flexible Discriminant Analysis by Optimal Scoring[J]. Publications of the American Statistical Association, 1994, 89(428): 1255–1 270.
[28] FRIEDMAN J H. Multivariate Adaptive Regression Splines[J]. The Annals of Statistics, 1991, 19(1): 1–67.
[29] PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions [J]. Ecological Modelling, 2006, 190(3): 231–259.
[30] SINGH K, MCCLEAN C J, BÜKER P, et al. Mapping regional risks from climate change for rainfed rice cultivation in India[J]. Agricultural Systems, 2017, 156: 76–84.
[31] 褚建民, 李毅夫, 张雷, 等. 濒危物种长柄扁桃的潜在分布与保护策略[J]. 生物多样性, 2017, 25(8): 799–806.
[32] ALLOUCHE O, TSOAR A, KADMON R. Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS) [J]. Journal of Applied Ecology, 2006, 43(6): 1223–1232.
[33] ANDERSON M J. Analysis of Ecological Communities[J]. Journal of Experimental Marine Biology & Ecology, 2003, 289.
[34] GAMA M, CRESPO D, DOLBETH M, ANASTÁCIO P M. Ensemble forecasting ofworldwide distribution: Projections of the impact of climate change[J]. Aquatic Conservation Marine & Freshwater Ecosystems, 2017, 27.
[35] 崔相艳, 王文娟, 杨小强, 李述, 秦声远, 戎俊. 基于生态位模型预测野生油茶的潜在分布[J]. 生物多样性, 2016, 24(10): 1117–1128.
[36] LIMA J S, BALLESTEROS-MEJIA L, LIMA-RIBEIRO M S, COLLEVATTI R G. Climatic changes can drive the loss of genetic diversity in a Neotropical savanna tree species[J]. Global Change Biology, 2017, 23(11): 4639.
[37] BROWN J L, YODER A D. Shifting ranges and conservation challenges for lemurs in the face of climate change[J]. Ecology & Evolution, 2015, 5(6): 1131–1142.
[38] 张成安, 丁长青. 中国鸡形目鸟类的分布格局[J]. 动物分类学报, 2008, 33(2): 317–323.
[39] WAN Jizhong, WANG Chunjing, TAN Jingfang, YU Feihai. Climatic niche divergence and habitat suitability of eight alien invasive weeds in China under climate change[J]. Ecology & Evolution, 2017, 7(5): 1541–1552.
[40] YAN Yujing, LI Yi, WANG Wenjing, et al. Range shifts in response to climate change of,, a fungus endemic to the Tibetan Plateau[J]. Biological Conservation, 2017, 206:143–150.
[41] ZHOU Wenbin, JI Xiang, OBATA S, et al. Resolving relationships and phylogeographic history of the, Nyssa sylvatica, complex using data from RAD-seq and species distribution modeling[J]. Molecular Phylogenetics and Evolution, 201, 126: 1–16.
Prediction ofspecies distribution based on multi-model combination
CHEN Sisi, LIU Xiang, TONG Xinyue, GUAN Bicai*
College of Life Sciences, Nanchang University, Nanchang 330031, China
Climate is a major factor affecting biodiversity and species distribution. In recent years, due to the large-scale emission of greenhouse gases, the climate change has a great impact on the distribution of plants. In this study,was used as the research object. Thedata and access of this field were used to obtain the distribution data. The environmental variables were obtained from the World Climate Website and Cold and Arid Regions Science Data Center at Lanzhou. The software of R, erdas, and ArcGIS were used to simulate the current and future (2070) distributions and derive confidence and calculate their area and centroid changes. The results show that all of the AUC values are greater than 0.9, which is a very good prediction. The main factors affecting the distribution are Bio01 (Mean annual temperature), Bio04 (Temperature seasonality), Bio15 (Coefficient of variation of Precipitation seasonality) and Bio03 (Isothermality and annual temperature difference ratio). In four kind of different CO2emission scenarios(RCP2.6, RCP4.5, RCP6.0 and RCP8.5), the result suggest that the distribution area ofHance. has contracted, and the shrinkage area increases with the increase of carbon emission. The maximum shrinkage area accounts for 80% of the total area. The survival of tea may be very threatening. In addition, under the background of the shrinkage of the total area, its distribution also appears in a trend of moving toward the north as a whole.
;biomod2; species distribution model; area change; centroid changes
10.14108/j.cnki.1008-8873.2020.02.008
Q948.13
A
1008-8873(2020)02-058-9
2018-10-11;
2010-02-01
国家自然科学基金(31360045); 江西省青年科学基金(20161BAB214160); 江西省研究生创新专项资金项目(YC2017-S038)。
陈思斯(1994—), 女, 硕士研究生, 研究方向为景观遗传学, E-mail: 513653721@qq.com
管毕财, 男, 博士, 副教授, 主要从事景观遗传学研究, E-mail:guanbicai12@163.com
陈思斯, 刘想, 童鑫玥, 等. 基于多模型集合预测尖萼红山茶物种分布[J]. 生态科学, 2020, 39(2): 58-66.
CHEN Sisi, LIU Xiang, TONG Xinyue, GUAN Bicai.Prediction ofspecies distribution based on multi-model combination[J]. Ecological Science, 2020, 39(2): 58-66.
