温敏核不育水稻HD9802-9S育性相关基因的BSA测序初步分析
2020-04-08甘丹阳李龙盘王倩徐国成居超明
甘丹阳,李龙盘,王倩,徐国成,居超明
(湖北大学生命科学学院,湖北 武汉 430062)
0 引言
温敏核不育(thermo-sensitive male sterile,TGMS)水稻属于环境敏感型不育系(environment-sensitive male sterile,EGMS).自1973年石明松发现光敏不育水稻NK58S后[1],EGMS开始应用于杂交水稻育种中.EGMS是一类花粉育性受外界环境变化而改变的水稻品系,它们在花粉发育的敏感时期,如果外界环境达到临界条件以上时,其花粉表现出不育性状,如果外界环境在临界条件以下时,其花粉表现为可育性状[2].TGMS属于对温度敏感的EGMS,在其小孢子发育时期对温度敏感,处于这一时期的TGMS水稻,如果外界温度高于临界温度,会导致水稻花粉不育[3].
HD9802S是本实验室自主培育的一个优质TGMS,其花粉不育性受一对主效隐性核基因tms5控制.该不育系具有转育起点温度低,育性稳定(需日均温低于23 ℃处理5 d以上才会转育),品质优,熟期短等优点[4].HD9802-9S是以HD9802S为母本、以固优12为父本杂交选育的不育系,人工气候箱鉴定结果显示HD9802-9S的转育起始温度高于HD9802S,大田的可繁性明显好于HD9802S,我们预测HD9802-9S和HD9802S之间有差异基因调控它们的花粉育性.
群体分离分析法(bulked segregant analysis,BSA)是Michelmore于1991年根据建立近等基因系(near iso-genic lines,NIL)类似的原理提出来的[5].该分析方法是将分离群体中表现出极端性状的个体混合起来构建混池,通过分析比对不同混池间的差异序列,筛选与目的基因紧密连锁的分子标记以达到定位目标基因的目的.BSA重测序是一个有效、可靠的技术手段,广泛应用于水稻的基因定位.Han等通过BSA重测序,结合SSR和序列标签位点(STS)定位水稻抗白叶枯病基因[6];Tiwari等通过BSA重测序结合SNP芯片定位水稻耐盐基因座[7];Sun等通过BSA重测序结合二代测序(NGS)技术(Seq-BSA),定位了水稻的抗寒基因座[8].我们采用的BSA重测序技术是利用QTL-seq[9]方法,定位HD9802-9S的育性相关基因,对比前期得到的HD9802S不育基因的定位结果[10],筛选HD9802-9S和HD9802S之间调控花粉育性的差异基因.
1 材料与方法
1.1 试验材料温敏核不育系HD9802-9S,恢复系R144和R446,HD9802-9S/R446 F2群体,HD9802-9S/R144 F2群体.
1.2 试验方法
1.2.1 花粉育性鉴定 两个F2代群体于2018年5月在武汉播种,6月插秧,每个群体数量约1 000株.七月底至八月中旬F2代群体处于抽穗期时对每一株材料进行花粉镜检并做标记.随机取抽穗期单株每穗上中下3个位置的花药置于滴有1%碘-碘化钾溶液的载玻片上,用镊子轻夹花药挤出花粉染色,在光学显微镜下放大160倍观察并记录.因为高温条件下,TGMS系不产生花粉,所以完全无花粉的单株标记为不育株,正常染色花粉超过80%的单株标记为高度可育株,如图1.
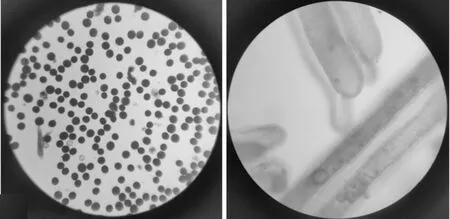
图1 高度可育株花粉镜检图(左),不育株花粉镜检图(右)
1.2.2 DNA样品制备 两个群体分别取高度可育株和不育株水稻叶片样本各100个,CTAB法抽提水稻基因组DNA,1%琼脂糖凝胶电泳检测DNA质量(若出现拖带,加入0.5 μL RNA酶A常温处理5~10 min).分别将HD9802-9S/R446 F2群体和HD9802-9S/R144 F2群体的不育株和高度可育株DNA等量混合构建HD9802-9S/R446 F2可育混池,HD9802-9S/R446 F2不育混池,HD9802-9S/R144 F2可育混池和HD9802-9S/R144 F2不育混池,获得混池样本,每个混池样本中DNA总含量为3~5 μg.用CTAB法分别抽提亲本DNA总量各3~5 μg构建亲本池,送武汉贝纳科技服务有限公司进行BSA重测序.
1.2.3 BSA重测序分析 以93-11的基因组序列为参考基因组,对所有混池进行二代测序,亲本池测序深度为60×,混池的测序深度为100×.通过计算两个子代样本相对于其中一亲本SNP位点的SNP-index值,计算两个极端混池样本之间的ΔSNP-index(ΔSNP-index = SNP-index(不育混池)-SNP-index(可育混池)),选取95%和99%置信水平作为筛选的阈值[9],以1 Mb为窗口,每次分析10 kb,统计窗口内SNP位点的ΔSNP-index平均值绘制ΔSNP-index沿染色体分布图.
2 结果与分析
2.1 BSA重测序的结果
2.1.1 HD9802-9S/R446 F2群体BSA重测序结果 HD9802-9S/R446 F2群体的ΔSNP-index沿染色体分布图如图2.以1 Mb为窗口,95%置信水平下超过阈值的连续区域作为候选区域,候选区域范围如表1,99%置信水平的候选区域如表2.
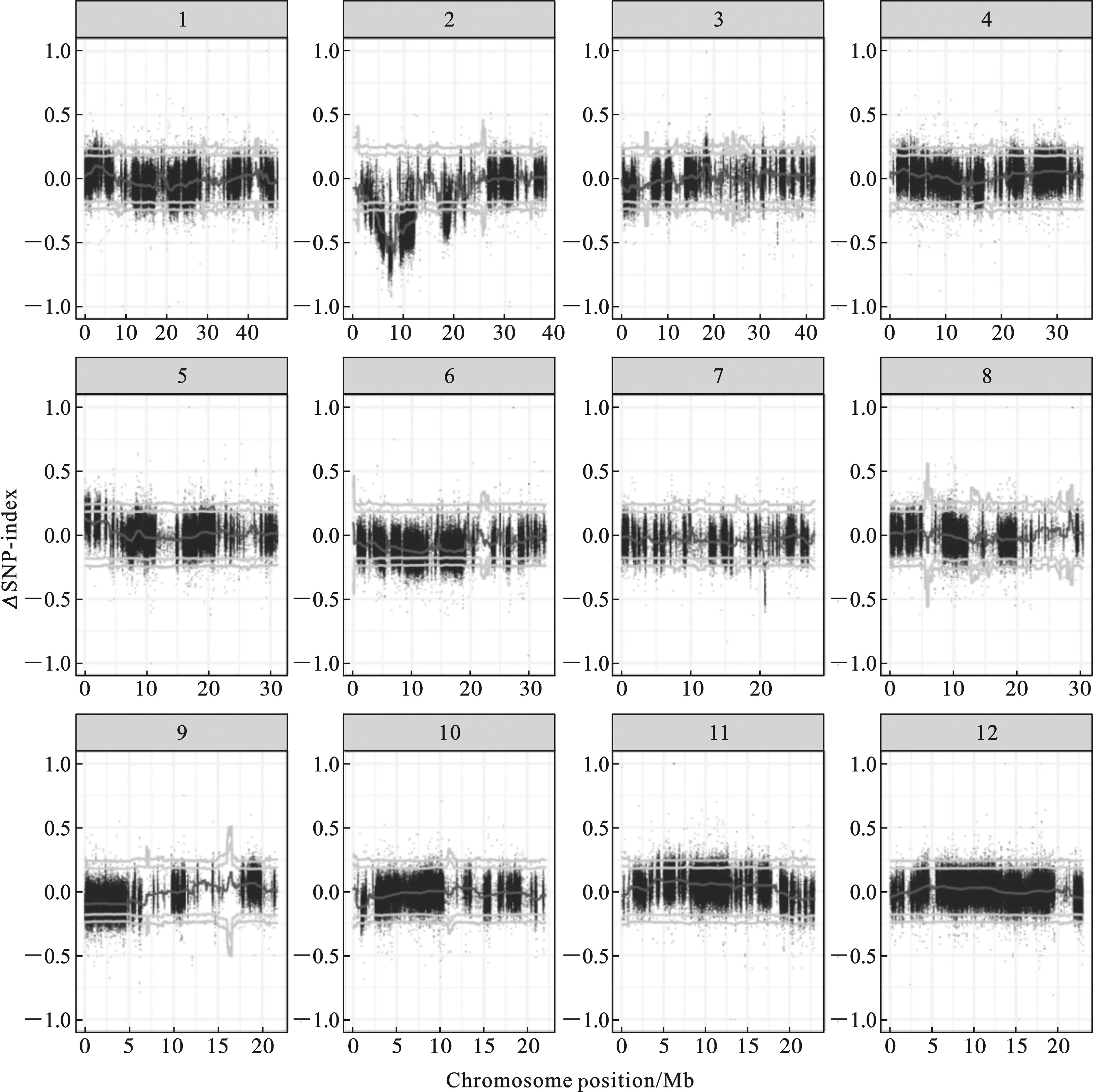
图2 HD9802-9S/R446 F2群体的ΔSNP-index沿染色体分布图
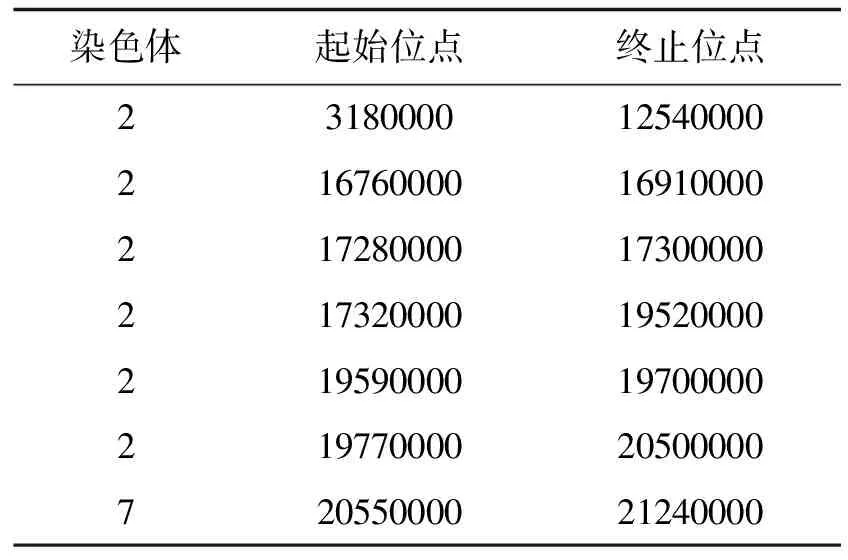
表1 HD9802-9S/R446 F2群体ΔSNP-index超过95%置信水平候选区域

表2 HD9802-9S/R446 F2群体ΔSNP-index超过99%置信水平候选区域
由图2和表1可以看出,当置信水平为95%时,HD9802-9S/R446 F2的ΔSNP-index集中在2号染色体上总长为12.57 Mb的候选区域内.且在7号染色体上有一段长为0.69 Mb的候选区域.当置信水平为99%时,HD9802-9S/R446 F2的ΔSNP-index集中在2号染色体上总长为9.3 Mb的候选区域内,在7号染色体上有一段长为0.16 Mb的候选区域,如表2,但在7号染色体这个区域内没有导致错译的SNP突变,由此推测7号染色体上的候选基因是影响HD9802-9S育性的微效基因.
2.1.2 HD9802-9S/R144 F2群体BSA结果 HD9802-9S/R144 F2群体的ΔSNP-index沿染色体分布图如图3,其置信水平95%的候选区域见表3,置信水平99%的候选区域见表4.
由图3和表3可以看出,当置信水平为95%时,HD9802-9S/R144 F2群体的ΔSNP-index集中在2号染色体上总长为3.24 Mb 的候选区域内,并且在7号染色体也上有一段长为0.69 Mb的候选区域.说明HD9802-9S的7号染色体上确实存在与水稻不育性状相关的突变基因.当置信水平为99%时,HD9802-9S的ΔSNP-index集中在2号染色体上总长为1.87 Mb的区域内,在7号染色体上有一段长为0.15 Mb的候选区域,如表4,但是这个候选区域内没有导致错译的SNP突变.
HD9802-9S/R144 F2和HD9802-9S/R446 F2两个群体的遗传背景是不相同的,但是通过BSA重测序得出的结果是相同的,与HD9802-9S育性相关的基因定位在2号染色体和7号染色体上.因为调控HD9802S育性的主效基因tms5定位于2号染色体短臂上[12],而HD9802-9S的不育基因来源于HD9802S,所以HD9802-9S调控育性的主效基因定位于2号染色体上,7号染色体上定位的候选基因为微效基因.

图3 HD9802-9S/R144 F2群体的ΔSNP-index沿染色体分布图
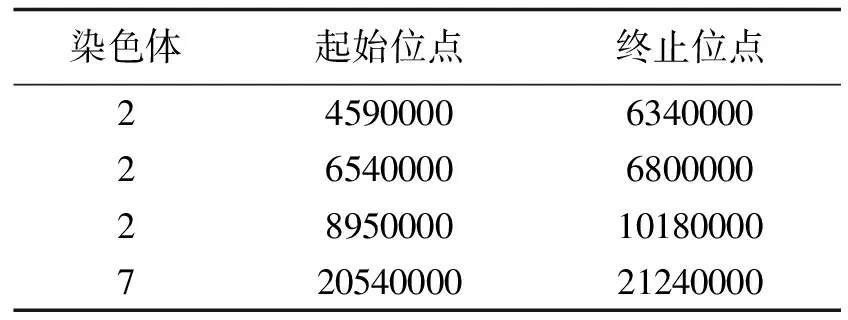
表3 HD9802-9S/R144 F2群体ΔSNP-index超过95%置信水平候选区域

表4 HD9802-9S/R144 F2群体ΔSNP-index超过99%置信水平候选区域
2.2 BSA重测序的数据分析HD9802-9S/R446F2群体的BSA重测序结果显示,置信水平为95%时,2号染色体上的候选区域上有1988个导致错义突变的SNP突变位点,覆盖431个候选基因,7号染色体上有15个导致错义突变的SNP突变位点,覆盖4个候选基因.置信水平为99%时,2号染色体上的候选区域上有1 423个导致错义突变的SNP突变位点,覆盖361个候选基因,7号染色体上没有导致错义突变的SNP突变位点(表5).HD9802-9S/R144 F2群体的BSA重测序结果显示,置信水平为95%时,2号染色体上的候选区域上有563个导致错义突变的SNP突变位点,覆盖120个候选基因,7号染色体上的候选区域上有15个导致错义突变的SNP突变位点,覆盖4个候选基因.置信水平为99%时,2号染色体上的候选区域上有367个导致错义突变的SNP突变位点,覆盖64个候选基因,7号染色体上没有导致错义突变的SNP突变位点(表6).

表5 HD9802-9S/R446F2群体SNP数量统计

表6 HD9802-9S/R144F2群体SNP数量统计
总SNP数量为95%水平的所有SNP数量,包含外显子上导致错译和不导致错译的单碱基替换和内含子上的单碱基突变,有效SNP数量为95%置信水平内导致错译的SNP数量
HD9802-9S/R446 F2和HD9802-9S/R144 F2两个群体的BSA重测序结果中,置信水平为95%时,两个群体重叠的候选基因基因共有88个,其中84个位于2号染色体上,4个位于7号染色体上.置信水平为99%时,两个群体重叠的候选基因共有64个,全部位于2号染色体上,7号染色体上没有候选基因.
置信水平为95%时,HD9802-9S的7号染色上的4个候选基因分别是BGIOSGA024073、BGIOSGA025960、BGIOSGA024051和BGIOSGA024043.在GO数据库(http://geneontology.org/)上查找这4个基因的注释,其中BGIOSGA024073基因的产物能够与ATP,核苷酸和未折叠的蛋白质结合;BGIOSGA025960基因的产物能够与钙离子结合,且功能与物质的跨膜转运相关;BGIOSGA024051基因与调控植物的昼夜节律相关;BGIOSGA024043基因还没有被注释.目前还没有这4个基因功能研究的相关报道.
置信水平为99%时,HD9802-9S的2号染色体上有64个候选基因,其中有44个基因在GO数据库上有注释.BGIOSGA006998、BGIOSGA007885、BGIOSGA006793、BGIOSGA006791、BGIOSGA006788、BGIOSGA007888、BGIOSGA006760、BGIOSGA006749、BGIOSGA007908基因产物是膜的组成部分;BGIOSGA007667基因产物是叶绿体被膜的组成部分;BGIOSGA007008基因参与调节翻译保真度;BGIOSGA007007基因产物有蛋白质-半胱氨酸S-棕榈酰转移酶活性;BGIOSGA006989基因与细胞核有关联;BGIOSGA007706、BGIOSGA006781、BGIOSGA006780基因产物可以与蛋白质结合;BGIOSGA006986、BGIOSGA006985、BGIOSGA006984参与跨膜转运过程;BGIOSGA006983基因产物与质膜有关;BGIOSGA006962基因参与GTP酶活性的正调节;BGIOSGA007886基因产物可以与序列特异性DNA结合;BGIOSGA006786、BGIOSGA006784、BGIOSGA007890、BGIOSGA007891与植物体内养分储存等活动相关;BGIOSGA007894基因参与调控植物根系发育;BGIOSGA007895基因产物有水解酶活性,可作用于酯键;BGIOSGA006775基因产物能够与核酸结合;BGIOSGA007898基因参与调控植物根冠发育;BGIOSGA006773基因参与转录调控,与DNA模板化相关;BGIOSGA006772基因参与植物的代谢过程;BGIOSGA006771、BGIOSGA006762基因产物具有蛋白质二聚活性;BGIOSGA006763基因产物可以与DNA结合;BGIOSGA006759基因参与植物对细菌的防御反应;BGIOSGA006758基因产物可以与锌离子结合;BGIOSGA006754基因参与调控植物对脱落酸的反应;BGIOSGA007905基因产物参与植物细胞壁组织组成;BGIOSGA006752、BGIOSGA006751基因产物具有电子载体活性;BGIOSGA006748基因与植物气孔复合体发育相关.还有20个基因没有注释.
3 讨论
我们分别构建HD9802-9S/R446和HD9802-9S/R144两个有不同父本遗传背景的F2代群体,将这两个群体的可育混池、不育混池和亲本池进行BSA重测序,把HD9802-9S的不育性状相关基因定位在2号染色体和7号染色体上.HD9802-9S的不育基因来源于HD9802S,而调控HD9802S的主效基因tms5的突变位点定位于2号染色体短臂上[11],所以HD9802-9S的BSA重测序结果中,超过99%置信水平的范围定位在2号染色体上是可信的.前期的研究中我们对HD9802S的不育基因做了定位分析[10],结果显示,在99%置信水平,HD9802S的不育相关基因主要定位在2号染色体上,在4号染色体上也存在超过99%置信水平的候选基因,在7号染色体上没有定位到候选基因.人工气候箱鉴定结果显示(表7,表8),HD9802S在14.5 h光照,23.5 ℃日均温条件下,花粉不育率为100%,对照培矮64S为99.24%±0.42%;HD98092-9S在14.5 h光照,24 ℃日均温条件下,花粉不育率为99.56%,对照培矮64S为99.48%.由此可以看出,HD9802-9S的转育温度比HD9802S的高.在海南连续三年冬季大田繁殖时,HD9802-9S的产量比HD9802S高,或者HD9802-9S产量底、而HD9802S基本绝收,说明HD9802-9S的转育启始温度高于HD9802S,或是HD9802-9S的低温耐受性弱于HD9802S.这进一步印证HD9802-9S和HD9802S的育性调控基因有差异,我们推测差异基因位于4号染色体和7号染色体上.
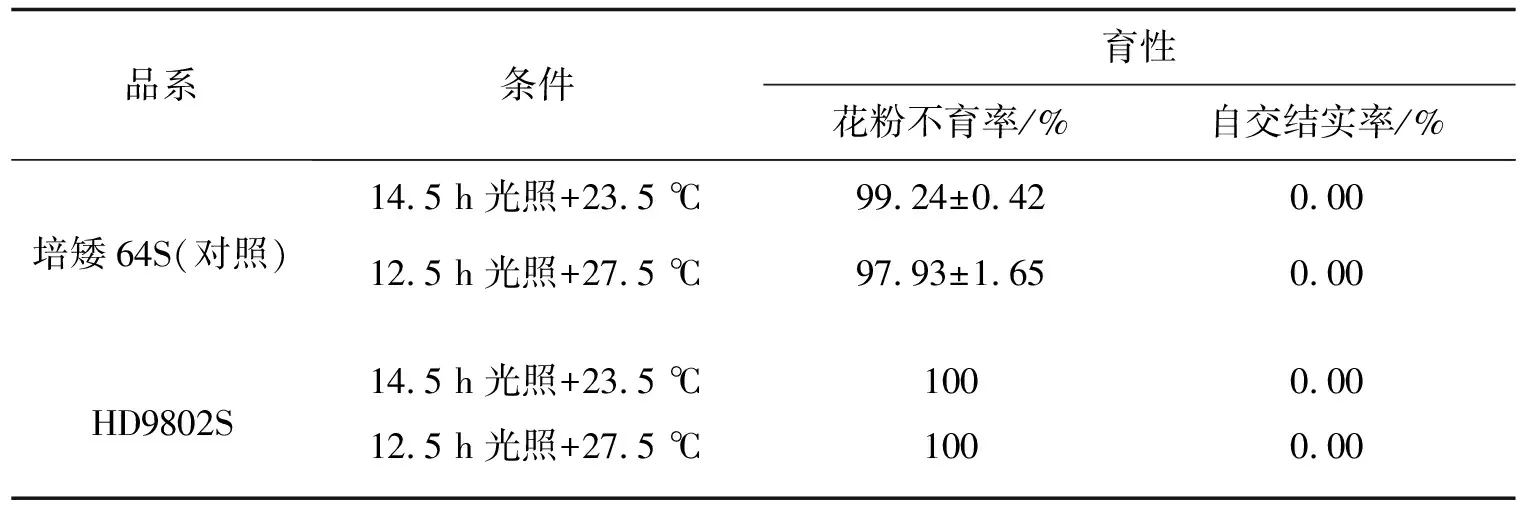
表7 HD9802S人工气候箱鉴定结果
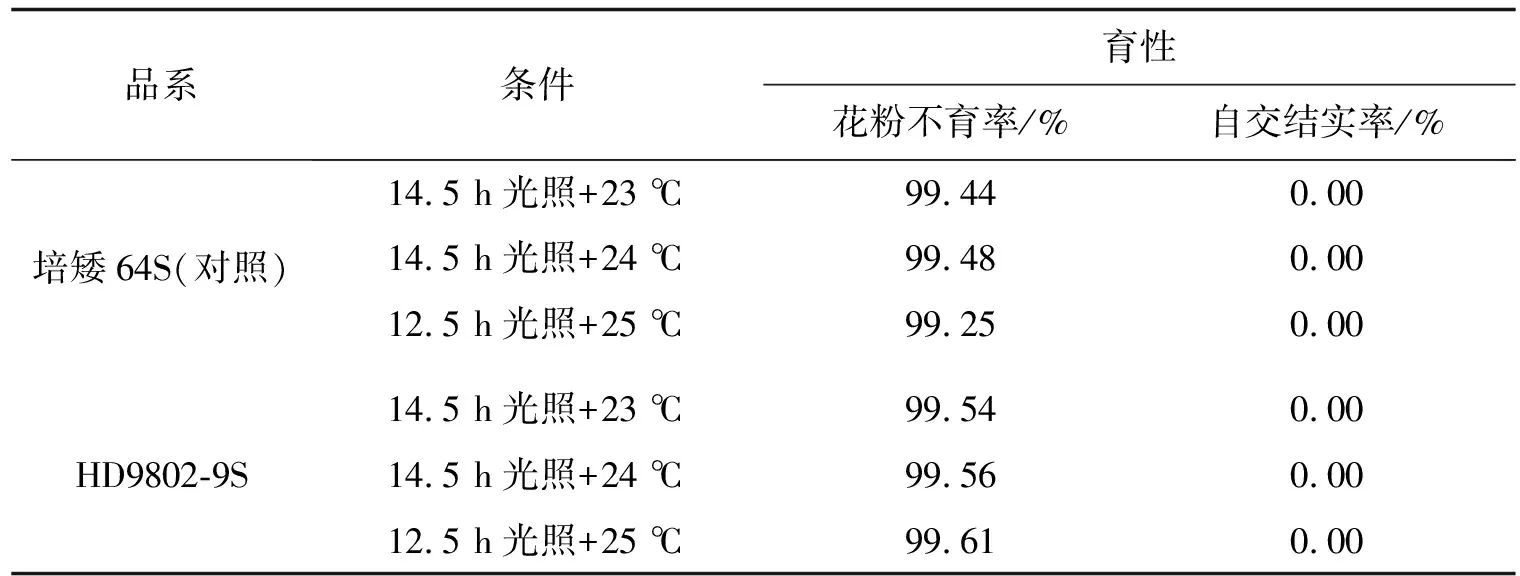
表8 HD9802-9S人工气候箱鉴定结果
tms5基因对水稻育性调控的分子机制已经被揭示.Zhou等人发现tms5基因的第71位碱基由C突变为A,产生一个提前终止的密码子,并将HD9802S的tms5基因进行测序发现HD9802S也存在这一突变,所以他们认为HD9802S的温敏不育性状的主效基因为tms5[11].这一结论与我们得出的BSA重测序结果一致.但是我们在HD9802S的4号染色体上定位并筛选出62个与花粉育性相关的候选基因,其中有20个候选基因位于99%置信水平,这62个候选基因中,有34个基因已经被注释.在HD9802-9S的7号染色体上也定位到4个候选基因,其中有3个基因已经被注释.这两个染色体上的候选基因中可能存在调控HD9802S和HD9802-9S之间花粉不育性差异的基因.在之后的研究中,我们主要研究定位于4号染色体和7号染色体上的候选基因,筛选出对tms5调控机制有影响、对水稻温敏不育性状有调控作用的微效基因,从而筛选出影响HD9802-9S和HD9802S花粉育性的差异基因,探究基因影响花粉育性的分子机制.
