干旱胁迫下杨树无性系苗期光合与气孔形态变异研究
2020-04-08乔滨杰王德秋高海燕李召珉葛丽丽丁文雅赵曦阳
乔滨杰 王德秋 高海燕 李召珉 葛丽丽 丁文雅 赵曦阳
(1.黑龙江省林业和草原局种苗总站,哈尔滨 150040; 2.临江林业局大西林场,临江 134600; 3.吉林省临江林业局,临江 134600; 4.东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040)
植物生长状况与环境因子密切相关,其中水分因素对植物影响较大[1]。在干旱、半干旱的恶劣条件下,水分的胁迫严重影响东北地区林木的生长[2],最终影响东北的木材生产及沙漠化防治。目前,针对杨树抗旱的研究与选育已经取得一定成果,高建社等[3]研究了5个杨树无性系冬季休眠期苗杆水分蒸腾速率、水分饱和亏缺、含水率,以及春季起苗后自然曝晒条件下苗木失水状况对造林后展叶时间的影响,并利用模糊数学隶属度公式对其抗旱性进行综合评价,选出84K杨为抗旱性最强的无性系。邱兴等[4]以6个杨树无性系为研究对象,通过人工盆栽控水试验,研究不同程度干旱胁迫对杨树无性系生长及生理性状的影响,通过隶属函数法进行综合评价,选出07-69×青1为抗旱性最强的杨树无性系。
光合作用是林木利用光能产生有机物和生长发育的重要基础[5]。叶片性状对环境变化最为敏感,在一定程度上叶片的变化反映了植物对资源摄取和利用的状态[6]。植物在受干旱胁迫影响后,其叶片含水量会发生显著变化,并对植物光合作用产生进一步影响[7]。干旱胁迫导致植物叶片光合速率降低、气孔阻力增大和二氧化碳交换速率下降,抑制了叶片的扩展及干物质的积累[8]。有研究表明水分胁迫下具有较高光合速率,且光合速率降低缓慢的植物具有较高的抗旱性[9]。因此对干旱胁迫下植物光合作用和叶片特征变化开展研究对植物抗旱性的鉴定具有重要意义。
杨树(Populusspp.)是我国人工林业产业化栽培的主要树种之一[10],在改善生态环境和解决木材短缺等方面具有重要地位[11]。但在干旱、半干旱地区,由于受到水分限制其生长受到抑制,严重影响了杨树在该地区的生态和经济效益[12]。目前已有部分关于杨树抗旱性的研究,主要集中在品种选育、生理生化研究和生长性状研究等方面,联合杨树幼苗的光合性状与气孔性状对无性系进行评价选择的研究较少,本研究对干旱胁迫下4个杨树无性系的叶片特征和光合作用进行测定,研究不同胁迫条件下杨树无性系的光合和叶片变化,选出抗旱性优良的无性系,为我国干旱、半干旱地区杨树抗旱无性系的评价选择提供材料和理论依据。
1 材料和方法
1.1 试验材料与处理
该试验于2018年4月在东北林业大学温室(126°61′E、45°78′N,海拔127 m)内开展,以白城小黑杨(Populussimonii×P.nigra‘Baicheng-1’)、白城小青黑杨(Populuspseudosimonii×P.nigra‘Baicheng-1’)、白林3号杨(Populussimonii×P.nigra‘Bailin-3’)及白城5号杨(Populus×Xiaozhanica‘Baicheng-5’)为材料进行扦插扩繁,每个无性系扩繁100株,放于高25.5 cm,口内径22.6 cm的花盆中。试验土壤配比:细沙∶黑土∶草炭土为2∶1∶1,每盆扦插1株,待成活后在温室内通过控水调节完成干旱胁迫试验。
采用室内环刀法[13]测定土壤田间持水量为28.90%。预实验于2018年6月下旬,每个无性系选择3盆长势一致的苗木作为预实验材料。开始开展预实验前后,对选出的各无性系苗木进行充分浇水,对浇水后的苗木进行命名、称重并详细记录。随后于每日上午8:30和下午5:00对预实验材料进行称重,记录每日土壤水分流失量,并观察苗木植株形态变化,当苗木叶片开始出现萎蔫时,增加称重次数至每日4次,对其重量变化进行详细记录并计算其土壤田间持水量,当叶片表现出大片枯死及脱落现象时,结束试验。通过预实验,总结出土壤田间持水量与浇水量之间的关系,以及如何通过浇水将土壤田间持水量保持在各干旱胁迫水平,根据预实验结果,试验设置1组对照和4种干旱胁迫水平,分别为土壤田间持水量的100%(对照)、80%、60%、40%和20%。预实验结束后,选取长势一致的各无性系苗木,每个无性系每种处理的选取10株苗木,共计200株。苗木选取完毕后通过称重法控制各处理土壤田间持水量至试验设计水平,于2018年7月中旬,正式开展试验,试验开展后每日进行1次称重,通过重量变化计算需补充水分使土壤水分维持在相应水平上。于每两天按照土壤田间持水量进行浇水一次。处理30 d后,各处理植株选取第5~7轮叶片进行光合作用参数和叶片气孔形态指标的测定。
1.2 各性状测定方法
利用Lico-6400光合测定系统进行光合作用参数的测定,测量时间为上午9:00~11:00,测定光照强度设定为1 000 μmol·m-2·s-1,CO2浓度设定为380 μmol·mol-1,测定叶片净光合速率(Net photosynthesis rate,Pn)、气孔导度(Stomatal conductance,Gs)、胞间CO2(Intercellular CO2dioxide concentration,Ci)和蒸腾速率(Transpiration rate,Tr),并计算水分利用效率(Water use efficiency):
WUE=Pn/Tr
(1)
气孔限制值(Limiting value of stomata):
Ls=1-Ci/Ca
(2)
式中:Ca为设定的CO2浓度,Ci为胞间CO2浓度。
气孔形态特征参数的测定采用指甲油印迹法。将叶片表面擦拭干净,选择叶片下表皮靠近叶脉部位均匀涂上无色指甲油,晾干,然后用透明胶带粘下指甲油薄膜层,剪成1 cm×1 cm方块,置于载玻片上展平,盖好盖玻片,用光学显微镜分别在100和400倍下观察并照相。在100倍镜视野下统计气孔数目,气孔密度=气孔数目/视野面积(Stomatal density,SD);在400倍镜视野下随机抽取完整清晰的气孔,测量气孔长度(Stomatal length,SL)、气孔器长度(Stomatal apparatus length,SAL)、气孔宽度(Stomatal width,SW),气孔器宽度(Stomatal apparatus width,SAW)、气孔面积(Stomatal area,SA)和气孔周长(Stomatal perimeter,SP)。每个叶片观测3个视野,共计测定30个视野。
1.3 数据统计分析
测定数据利用Excel 2016进行整理和作图,利用SPSS22.0进行数据方差分析,同时利用隶属函数评估法对4个无性系的光合特性和气孔形态指标进行抗旱性评价。
方差分析线性模型为:
Xijk=μ+Ci+Bj+CBij+eijk
(3)
式中:μ为总体平均值,Ci为无性系效应,Bj为处理效应,CBij为无性系和处理的交互效应值,eijk为环境误差。
表型变异系数[14]:
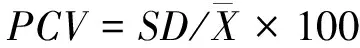
(4)
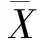
采用布雷津多性状综合评定法对无性系进行综合评定,具体公式为[12]:
(5)
式中:Qi为综合评价值,Xij为某一性状的平均值,Xjmax为某一性状的最优值,n为评价指标的个数。
表型相关分析采用公式[15]:
(6)
隶属函数法采用公式[16]:
当某一指标与林木抗旱性呈正相关采用公式:
U(Xi)=(Xi-Xmin)/(Xmax-Xmin)
(7)
当指标与林木抗旱性呈负相关,则采用反隶属函数进行转换,公式为:
U(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)
(8)
式中:U(Xi)为隶属函数值,Xi为干旱胁迫下指标测定值与正常水分下性状测定值的比值,Xmax、Xmin为所有参试品种某一指标相应比值的最大值和最小值。
2 结果与分析
2.1 方差分析
不同处理下各性状方差分析结果见表1,除气孔器长在处理间差异不显著外,其余各性状在无性系间、处理间和无性系与处理交互作用间均达到极显著差异水平。
2.2 不同干旱胁迫处理下杨树光合响应变化
由图1~2可知,随着土壤水分的逐渐减少,杨树光合作用受到明显的抑制,净光合速率和气孔导度均呈现持续下降趋势,其中白城小青黑杨在不同胁迫梯度上均优于其他无性系;当土壤相对含水量为60%时,白城小青黑杨、白林3号杨、白城5号杨和白城小黑杨的净光合速率分别为对照的88.29%、83.07%、74.66%和71.56%,与此同时,白城小青黑杨和白林3号杨气孔导度明显高于白城小黑杨和白城5号杨;当土壤田间持水量为40%时,白城小黑杨与白城5号杨净光合速率下降幅度加大,下降值分别为对照的63.40%和65.46%。当土壤田间持水量为20%时,各无性系光合值趋近于0,且各无性系气孔导度值相比于对照分别下降了79.63%(白城小黑杨)、80.33%(白林3号杨)、82.57%(白城5号杨)和80.69%(白城小青黑杨)。由图3~4可以看出,随着土壤相对含水量的逐渐降低,各无性系气孔限制值呈现先上升后下降的趋势,与之相反的,胞间二氧化碳浓度均呈现出先下降后上升的趋势;当土壤相对含水量高于60%时,胞间二氧化碳浓度呈下降趋势,当土壤相对含水量低于60%时,胞间二氧化碳浓度呈上升趋势。
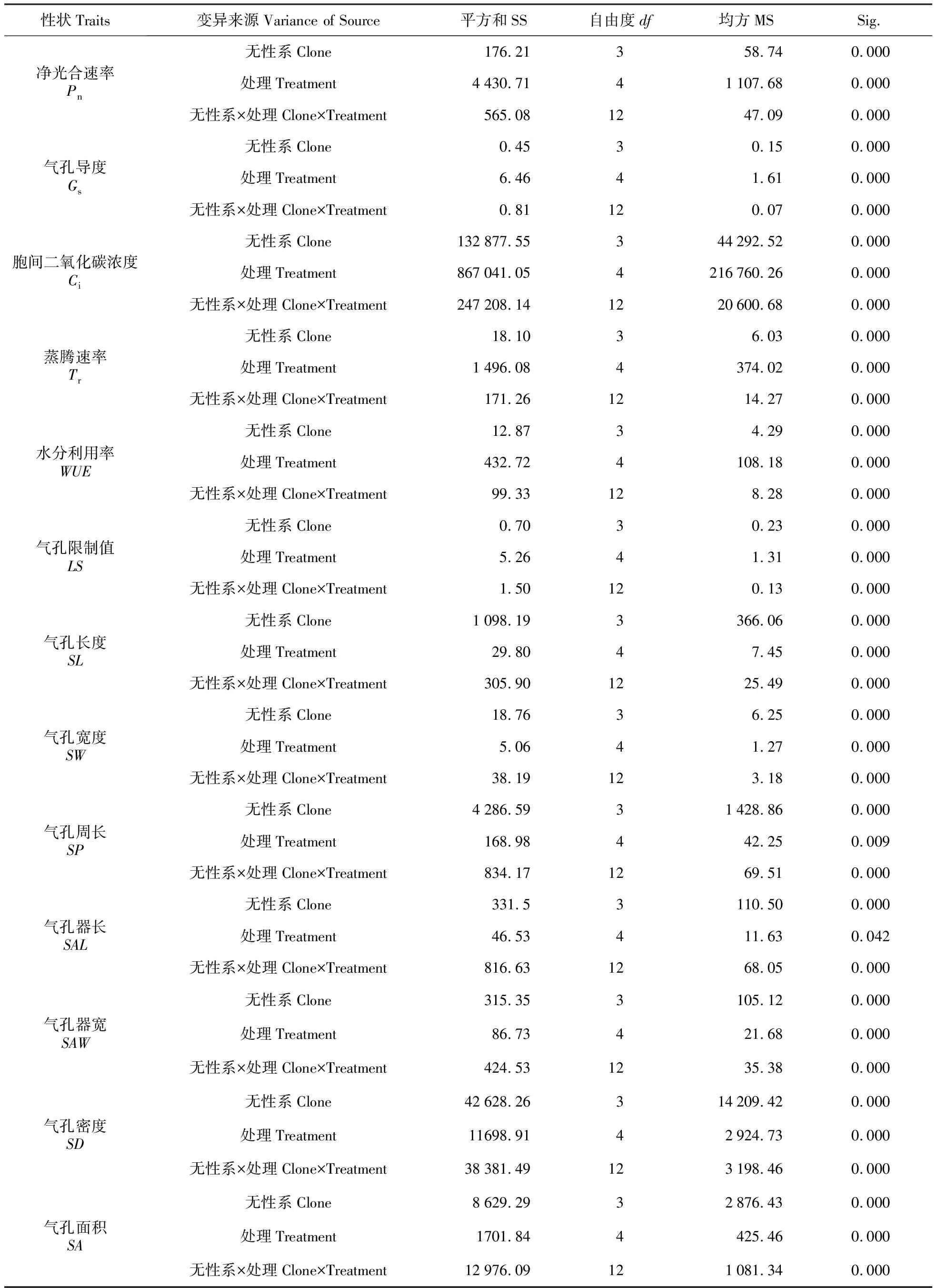
表1 干旱胁迫处理下各性状方差分析
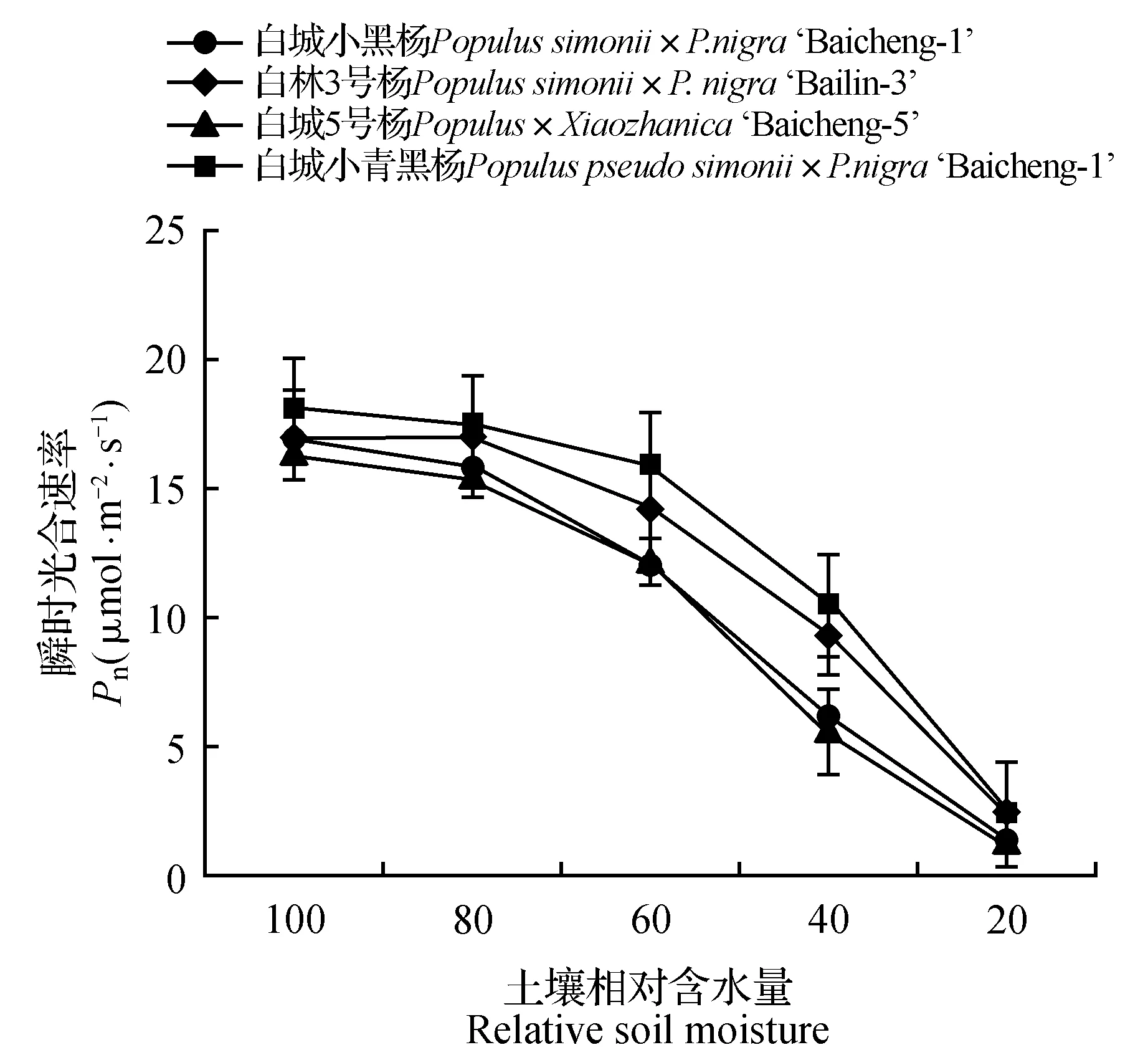
图1 干旱胁迫下叶片净光合速率变化Fig.1 Pn under different drought stress
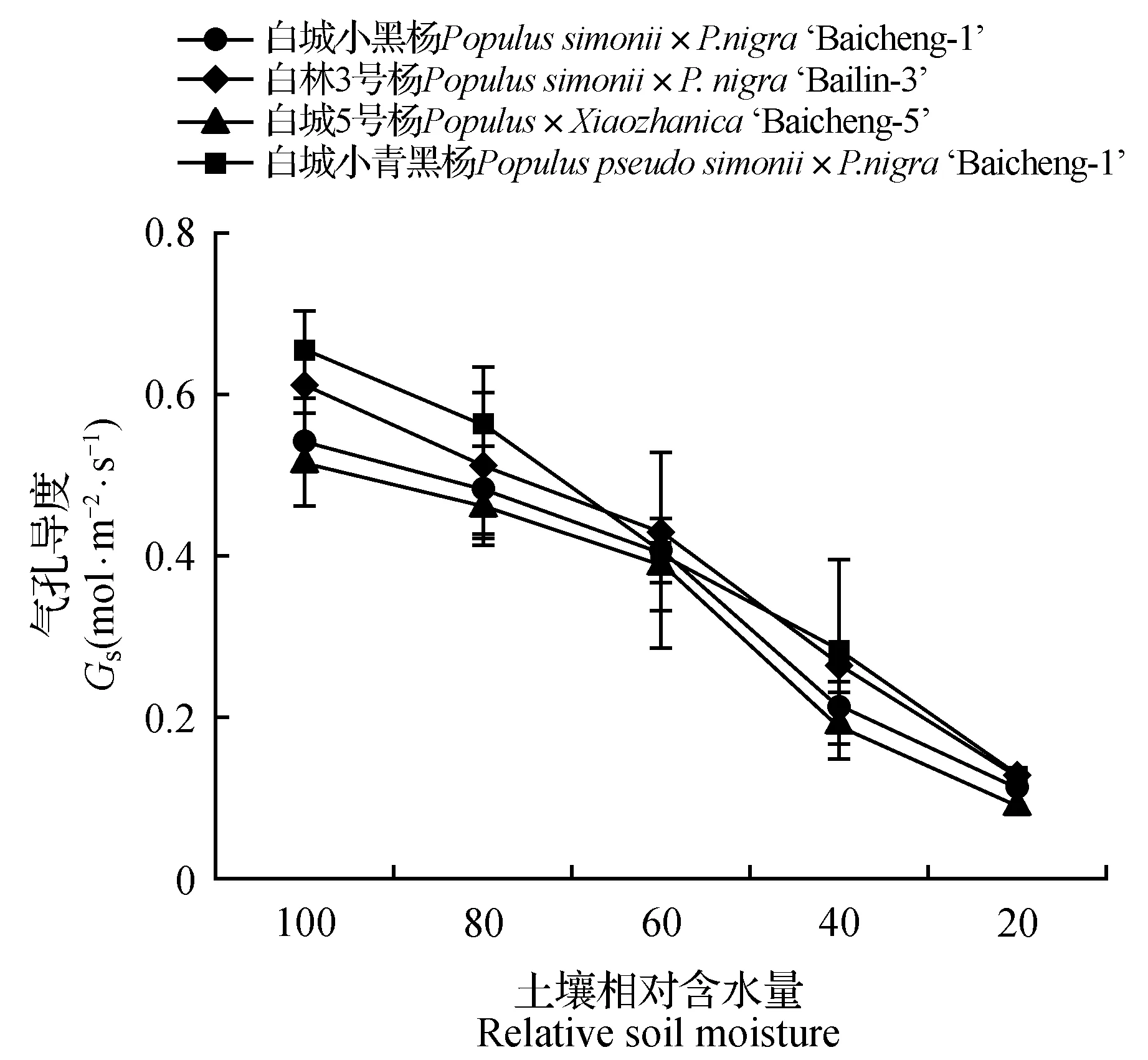
图2 干旱胁迫下叶片气孔导度变化Fig.2 Gs under different drought stress
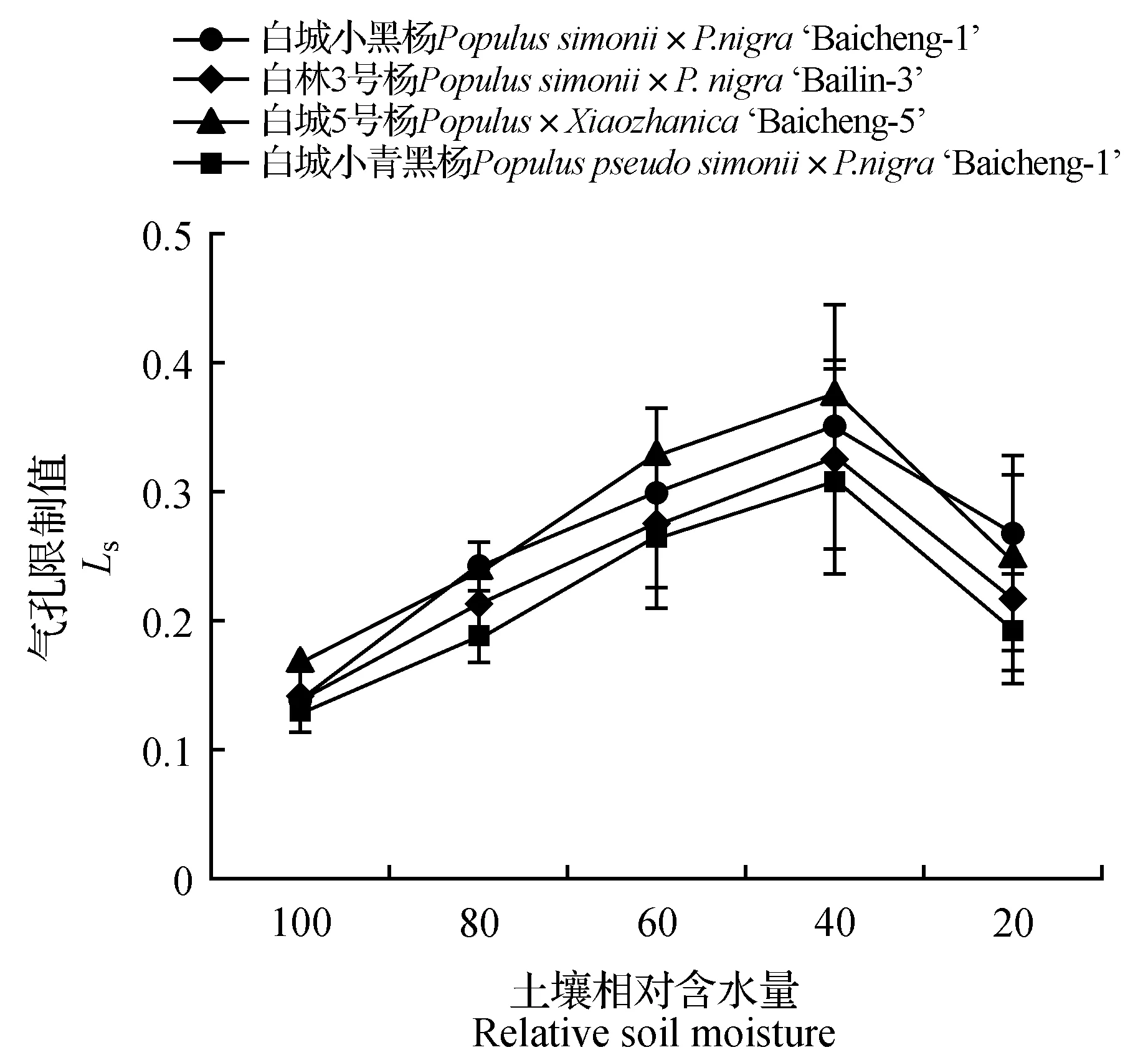
图3 不同干旱胁迫下叶片气孔限制值变化Fig.3 Ci/Ca under different drought stress
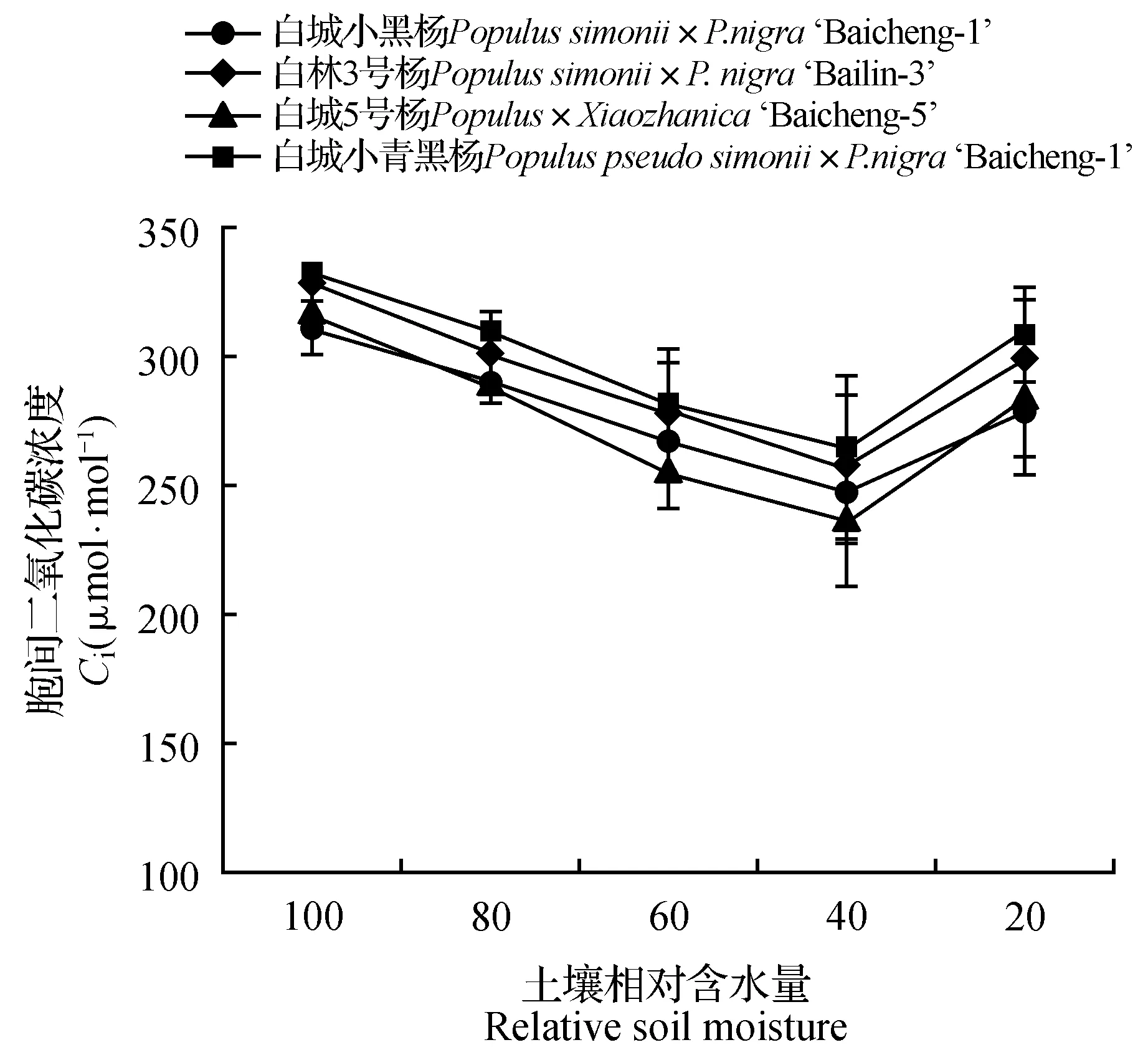
图4 不同干旱胁迫下叶片胞间二氧化碳浓度变化Fig.4 Ci under different drought stress
2.3 不同干旱胁迫处理下杨树叶片水分利用率和蒸腾速率变化规律
由图5可知,白城小青黑杨和白林3号杨对照处理的水分利用率较白城5号杨和白城小黑杨的高。随着干旱胁迫的加剧,除白城小黑杨在土壤相对含水量为80%时出现轻微下降外,各无性系水分利用率均呈现先上升后下降的趋势;当土壤田间持水量为60%时,各无性系水分利用率相比于对照分别上升了4.36%(白城小黑杨)、18.24%(白林3号杨)、12.52%(白城5号杨)和21.27%(白城小青黑杨);当土壤相对含水量低于60%时,各无性系水分利用率出现明显下降趋势,其中白城5号杨下降最为明显;当土壤田间持水量为20%时各无性系水分利用率相比于对照分别下降了44.52%(白城小黑杨)、24.08%(白林3号杨)、46.65%(白城5号杨)和26.54%(白城小青黑杨)。由图6可知,各无性系蒸腾速率随着干旱胁迫的加剧均呈现下降趋势;随着干旱胁迫程度的增大,各无性系均出现急剧下降,当土壤田间持水量下降为20%时,各无性系蒸腾速率分别为对照的15.25%、17.76%、15.30%和18.22%。
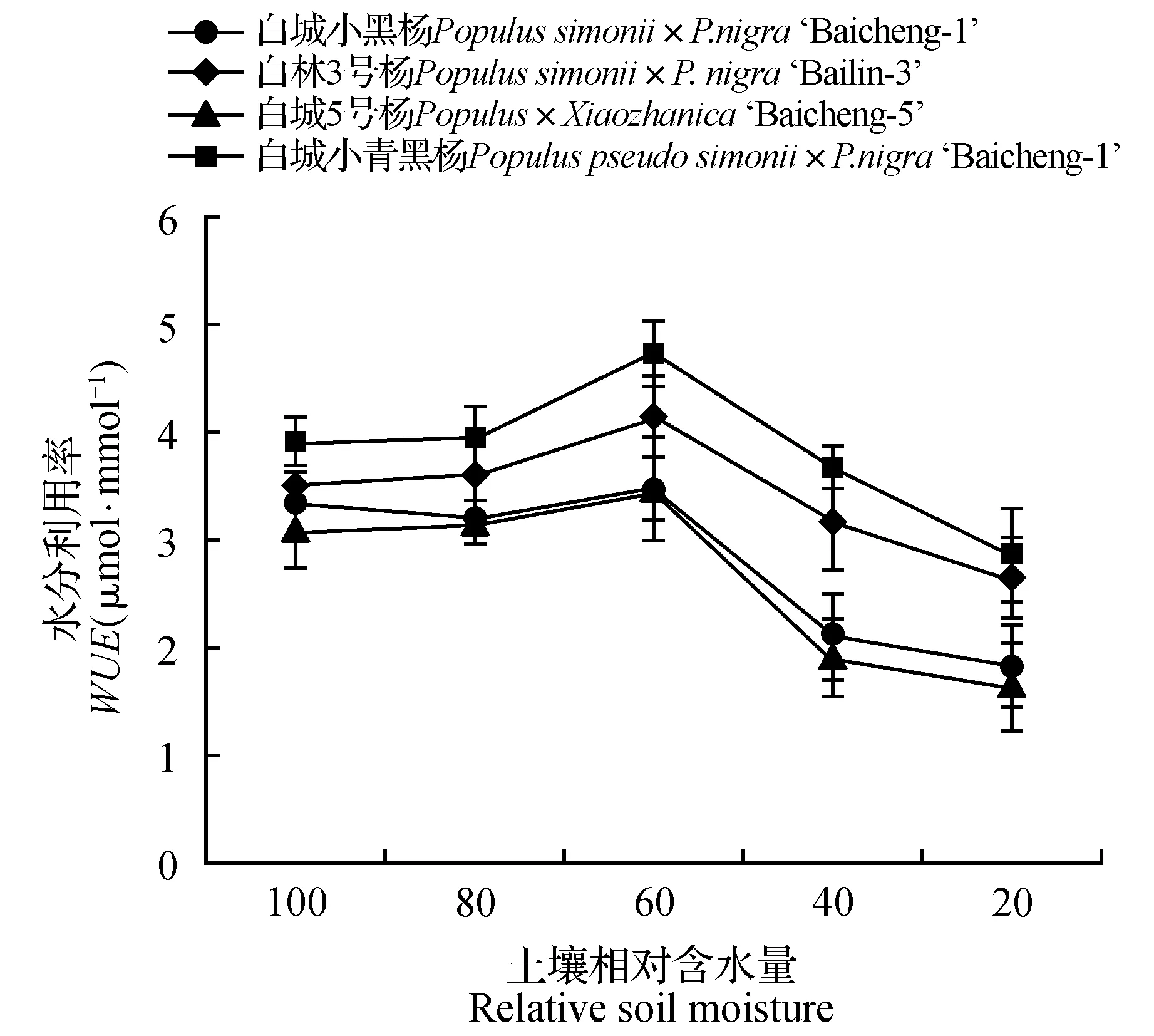
图5 不同干旱胁迫下叶片水分利用率变化Fig.5 Wue ueder different drought stress
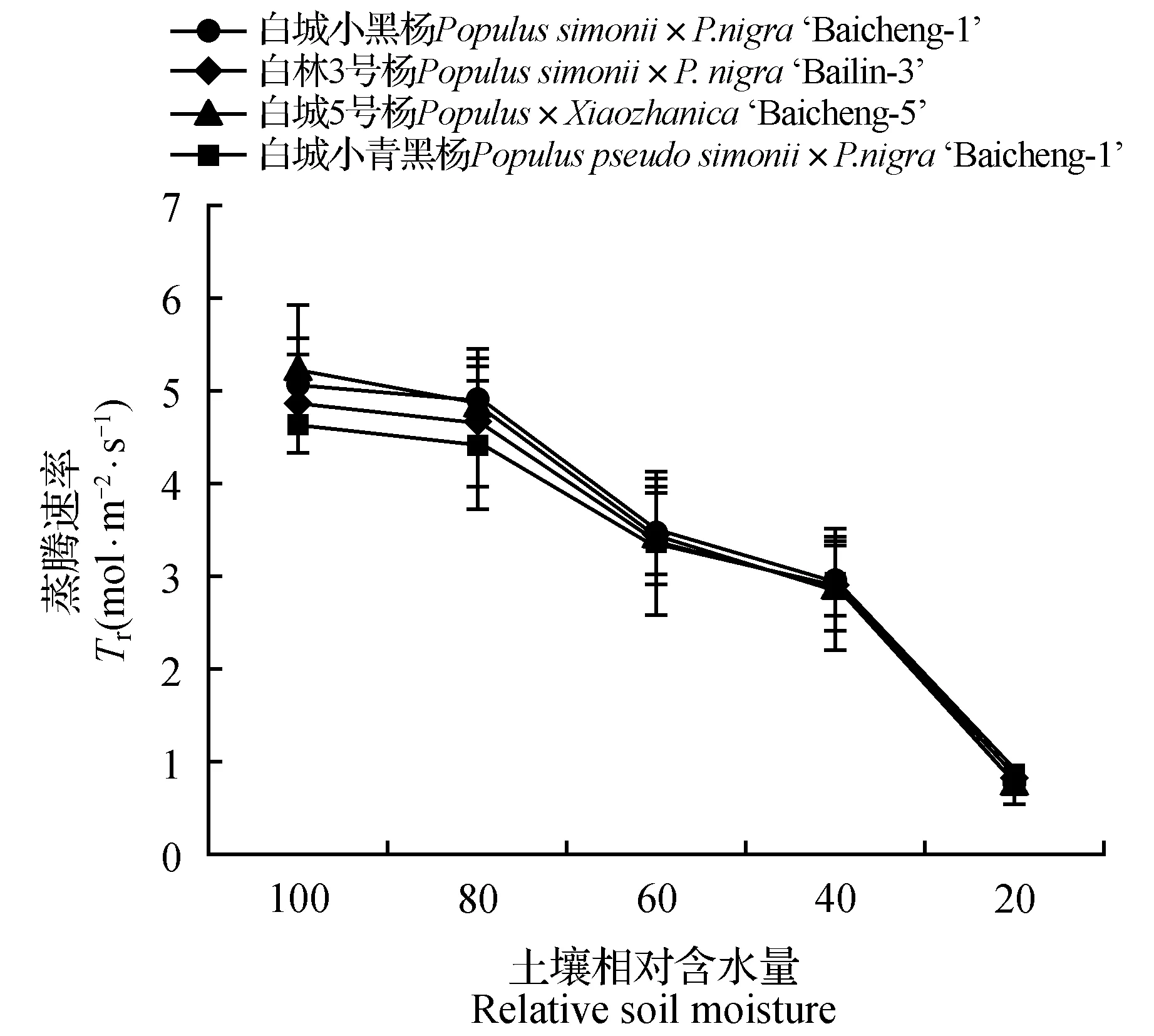
图6 不同干旱胁迫下叶片蒸腾速率变化Fig.6 Tr under different drought stress
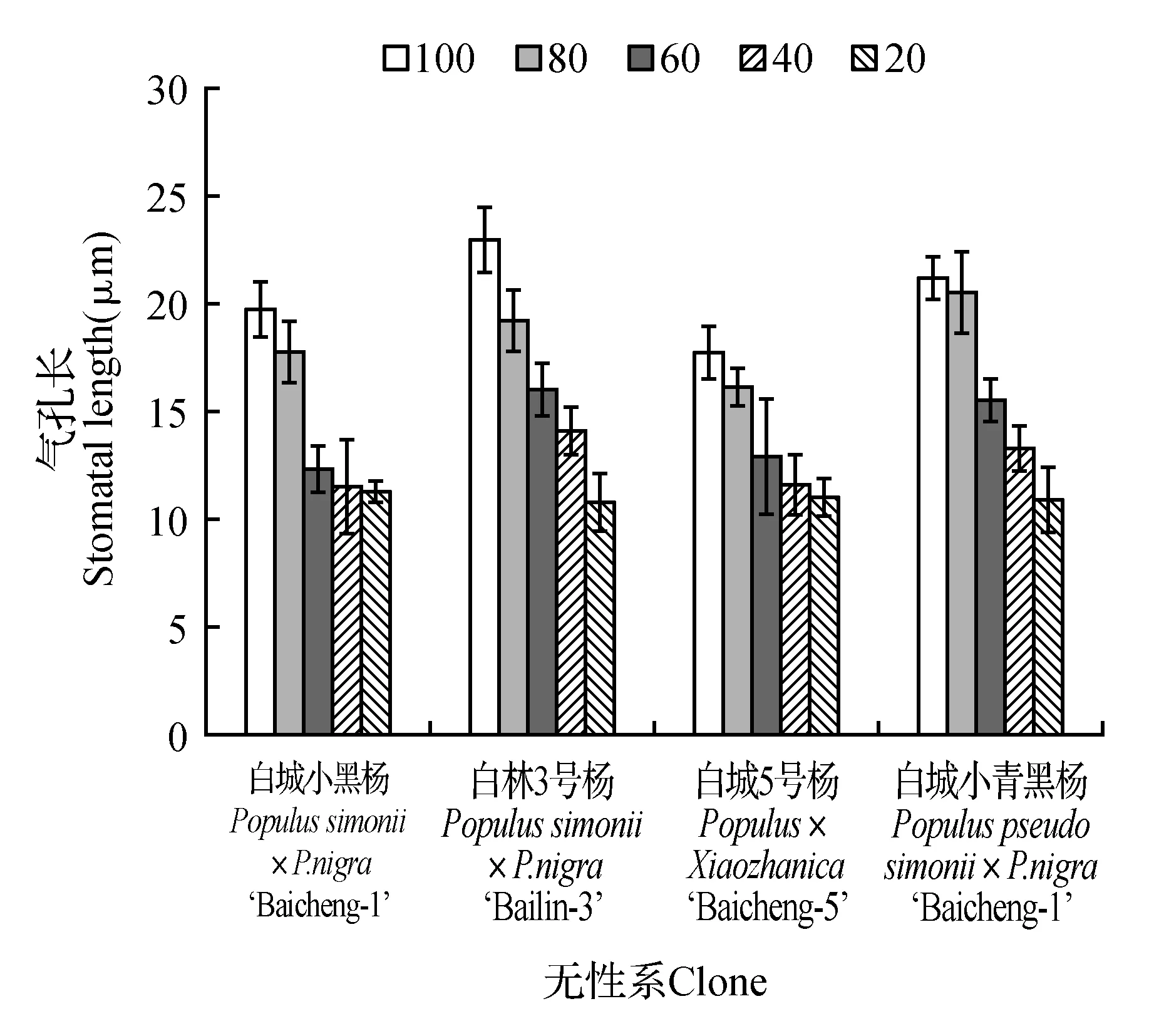
图7 不同干旱胁迫下叶片气孔长变化Fig.7 SL under different drought stress
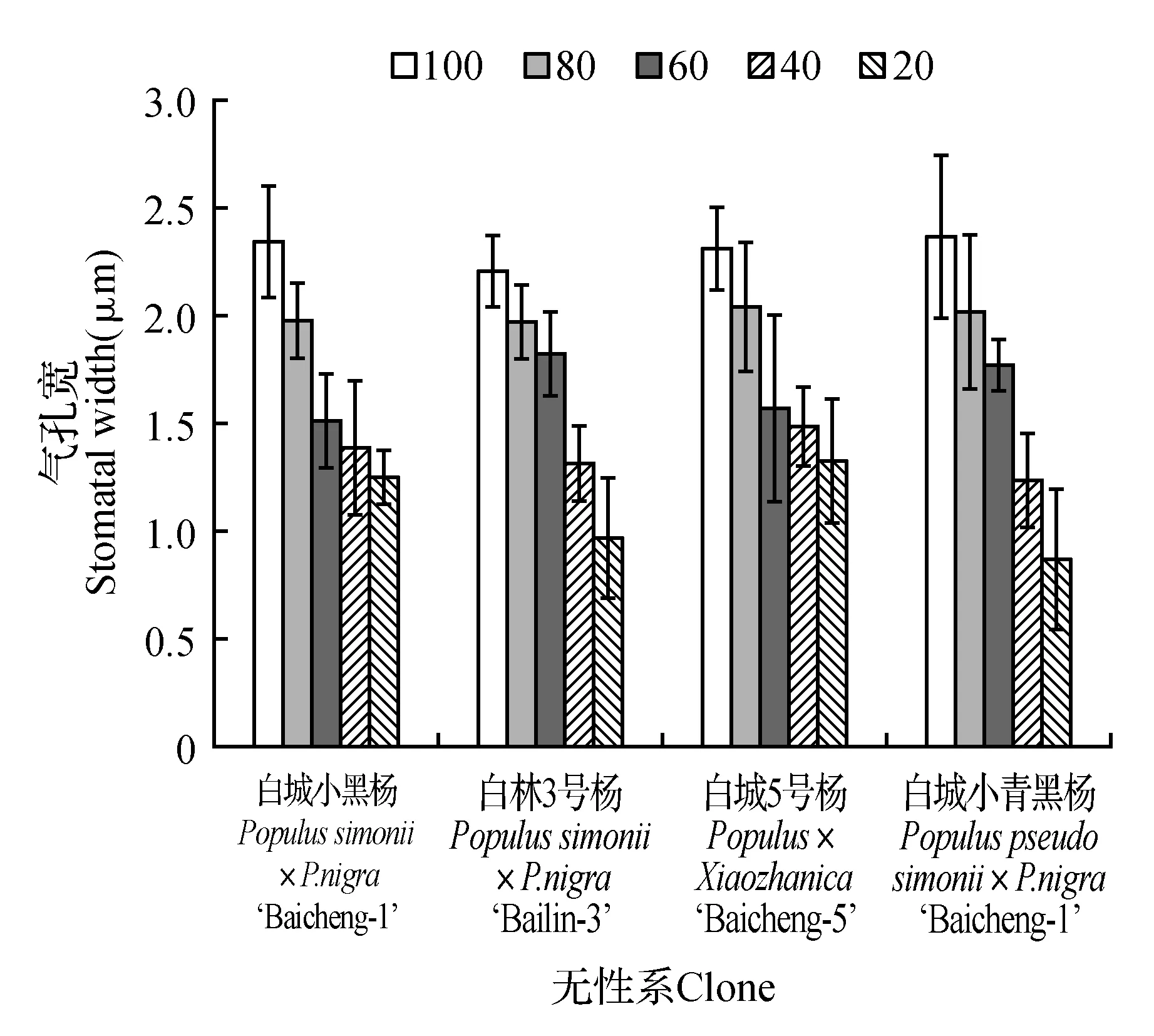
图8 不同干旱胁迫下叶片气孔宽变化Fig.8 SW under different drought stress

图9 不同干旱胁迫下叶片气孔周长变化Fig.9 SP under different drought stress
2.4 不同干旱胁迫处理下杨树叶片气孔形态变化规律
由图7可知,各无性系气孔长、气孔宽、气孔周长、气孔器长和气孔器宽均随着土壤相对含水量的降低呈现出不同程度的降低,但不同无性系间差异较大。由图7可知,当土壤相对含水量为60%时,各无性系均出现显著低于对照,分别降低为对照的62.30%(白城小黑杨)、69.86%(白林3号杨)、73.14%(白城5号杨)和73.25%(白城小青黑杨);随着土壤相对含水量的持续降低,白城5号杨和白城小黑杨的降低速度明显低于其他无性系。叶片气孔宽变化规律与叶片气孔长变化规律相似(图8),在土壤相对含水量为80%时,白城小黑杨下降幅度最大为对照的15.38%;当土壤田间持水量为60%时,白城小黑杨和白城5号杨下降最为明显,相比于对照分别降低了35.47%和31.60%;当土壤相对含水量为20%时,各无性系降至最低值,白城小黑杨、白林3号杨、白城5号杨和白城小青黑杨相比于对照分别降低46.88%,56.11%,42.86%和63.29%。各无性系不同处理下气孔周长变化如图9所示,各无性系均随土壤相对含水量的降低而减小,其中白林3号杨在土壤相对含水量为80%时,下降最为显著;四个杨树无性系在60%时均出现明显下降,降低至对照的63.89%(白城小黑杨)~75.06%(白城小青黑杨);当土壤相对含水量低于60%时,白城小黑杨和白城5号杨下降趋势减缓。
由图10可知,各无性系气孔器长、宽随着土壤相对含水量的降低,均表现为下降趋势。从气孔器长的变化趋势来看,除白城5号杨在80%处理下表现出明显降低,其余无性系在80%处理下与对照相差不明显;当土壤相对含水量为20%时各无性系均表现出最低值,分别为对照的74.19%(白城小黑杨)、75.93%(白林3号杨)、79.11%(白城5号杨)和73.99%(白城小青黑杨)。各无性系气孔器宽变化规律如图11所示,不同无性系变化趋势不同,其中白城小黑杨与白城5号杨变化幅度较其余2个无性系小;除白城小青黑杨60%处理与80%处理变化较小外,其余处理间变化较大;当土壤相对含水量为20%时各无性系相比于对照分别降低11.84%(白城小黑杨)、32.22%(白林3号杨)、7.27%(白城5号杨)和27.64%(白城小青黑杨)。
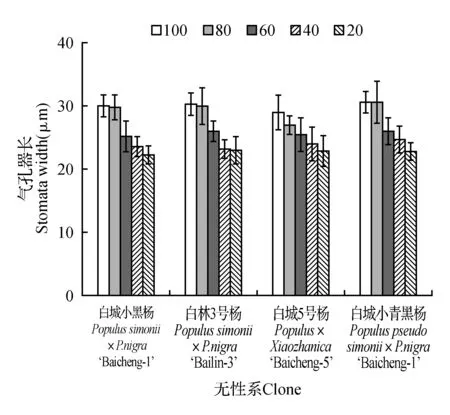
图10 不同干旱胁迫下叶片气孔器长变化Fig.10 SAL under different drought stress
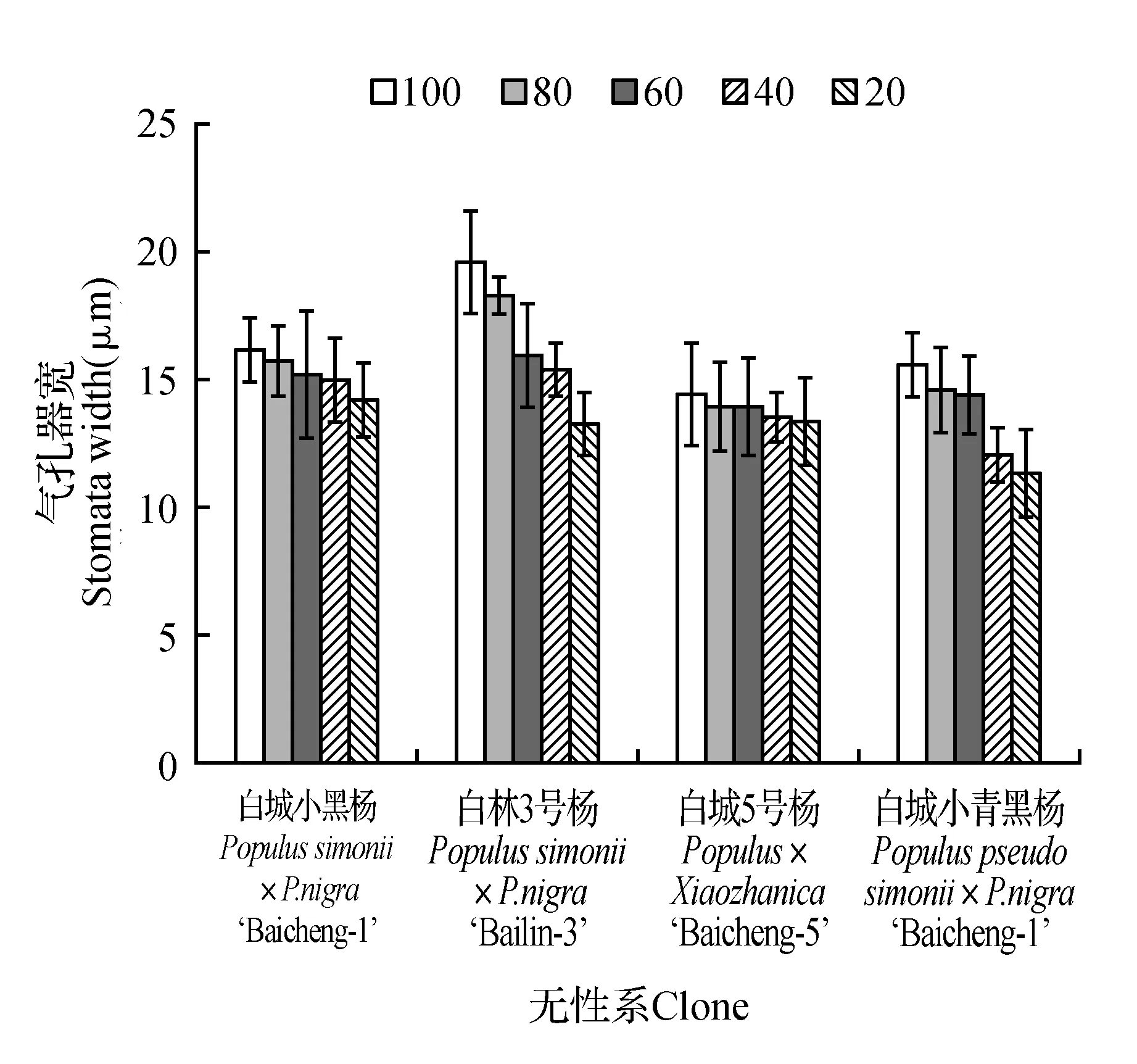
图11 不同干旱胁迫下叶片气孔器宽变化Fig.11 SAW under different drought stress
2.5 不同干旱胁迫处理下杨树叶片气孔密度与气孔面积的变化规律
由图12可以看出,4个杨树品种随着土壤相对含水量的降低,各无性系不同处理的叶片气孔密度呈现逐步下降,处理组和对照组的叶片气孔密度相比具有明显降低;随着土壤相对含水量的降低,各无性系呈现逐步下降的趋势。当土壤田间持水量为20%时,各无性系气孔密度降低至最小值,相比于不同无性系受胁迫影响气孔密度下降幅度不同,当土壤田间持水量为20%时,各无性系气孔密度降低至最小值,分别为对照组的73.32%(白城小黑杨)、77.68%(白林3号杨)、81.57%(白城5号杨)和77.60%(白城小青黑杨)。杨树叶片气孔面积随土壤相对含水量的变化如图13所示。随着土壤相对含水量的降低,各无性系均表现为持续下降趋势,不同土壤相对含水量下各无性系气孔面积最低值分别较各自对照组降低25.65%(白城小黑杨)、45.27%(白林3号杨)、35.58%(白城5号杨)和38.37%(白城小青黑杨)。
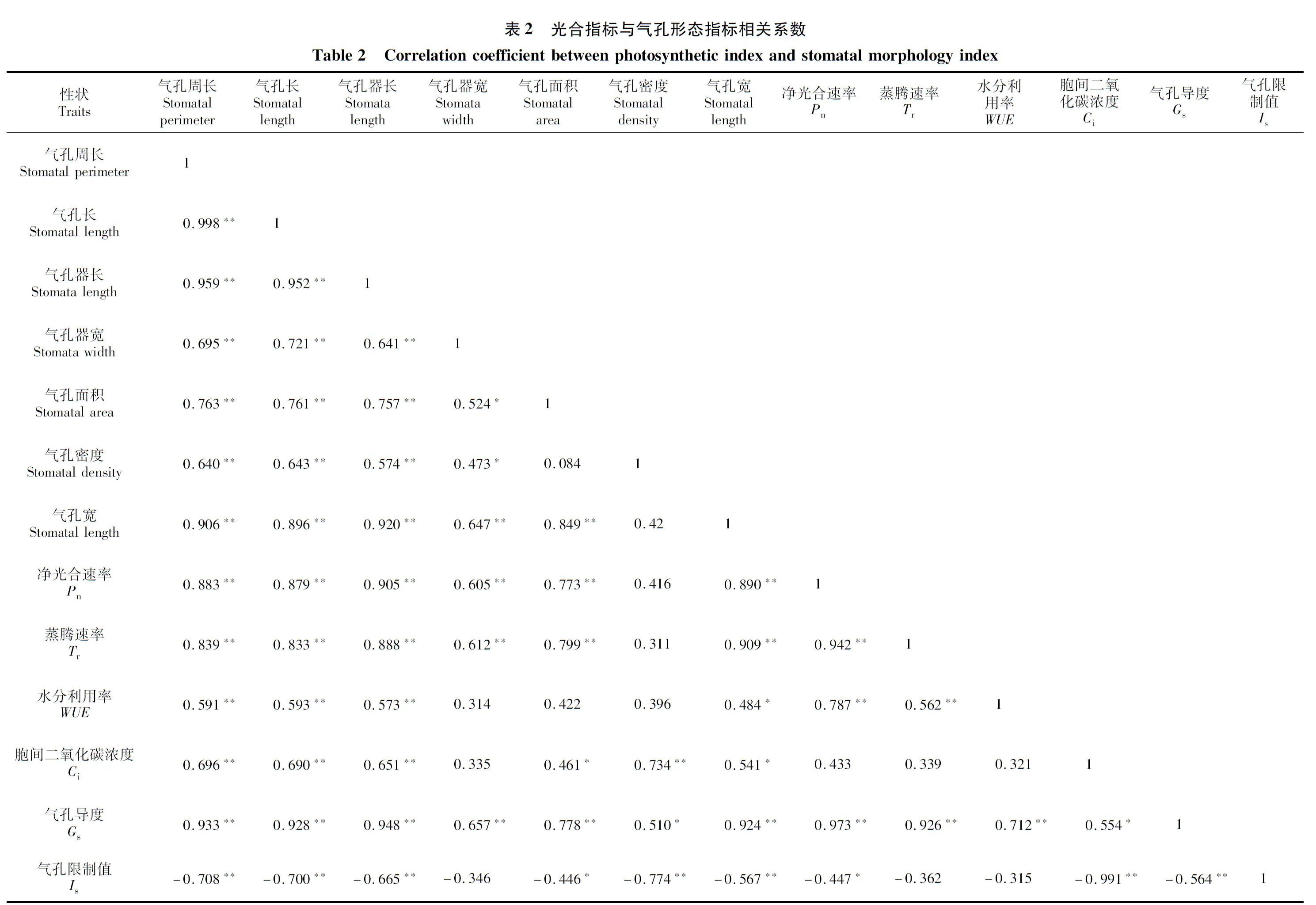
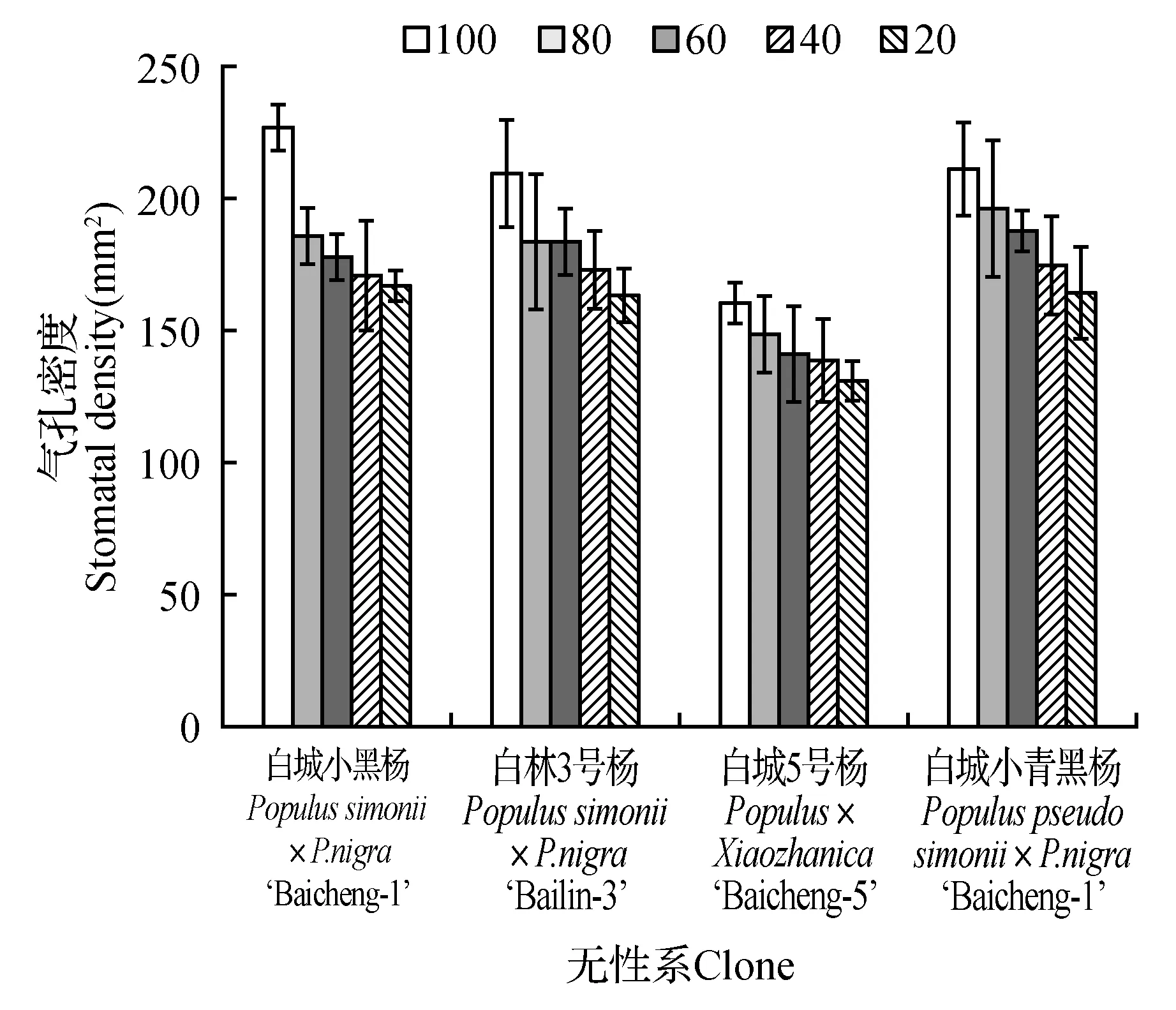
图12 不同干旱胁迫下叶片气孔密度变化Fig.12 SD under different drought stress
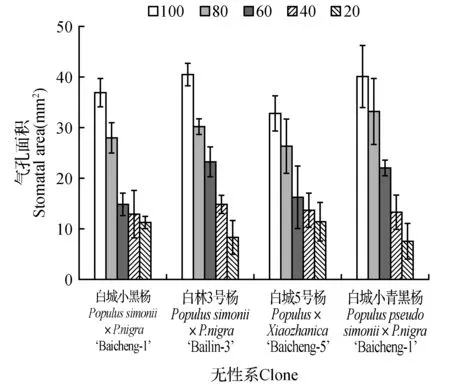
图13 不同干旱胁迫下叶片气孔面积变化Fig.13 SA under different drought stress
2.6 不同干旱胁迫处理下各指标相关性分析
各指标相关系数见表2,从气孔形态指标来看,除气孔器宽、气孔面积和气孔密度呈显著相关(0.524,0.473),气孔密度与气孔面积相关性无显著差异(0.084),其余指标间均达到极显著正相关水平(0.640~0.998)。对光合指标相关性分析发现净光合速率、蒸腾速率、水分利用率和气孔导度间呈极显著正相关关系(0.562~0.973),气孔限制值与胞间二氧化碳浓度和气孔导度呈极显著负相关关系(-0.991,-0.564)。对气孔形态指标与光和指标相关性分析发现,除气孔限制值外均表现为正相关关系(0.311~0.948),其中大部分指标间呈极显著正相关关系。
2.7 4个杨树无性系抗旱性的综合评价
利用隶属函数法对4个杨树无性系的抗旱性进行综合评价。由表3可知,当土壤相对含水量为80%时,白林3号杨、白城小青黑杨、白城小黑杨和白城5号杨的平均隶属函数值分别为0.51,0.66,0.55和0.46;当土壤相对含水量为60%时,白林3号杨、白城小青黑杨、白城小黑杨和白城5号杨的平均隶属函数值分别为0.57、0.72,0.36和0.53;当土壤相对含水量为40%时,白林3号杨、白城小青黑杨、白城小黑杨和白城5号杨的平均隶属函数值分别为0.51,0.67,0.41和0.46;当土壤相对含水量为20%时,白林3号杨、白城小青黑杨、白城小黑杨和白城5号杨的平均隶属函数值分别为0.44,0.51,0.47和0.50。综合分析表明,不同土壤相对含水量下白城小青黑抗旱性较高,白林3号杨次之。

表3 4个杨树无性系抗旱性的隶属函数分析
3 讨论
干旱是限制植物生长的重要非生物胁迫因素之一[17]。在干旱条件下生长时,植物的呼吸作用、光合作用和气孔作用等会受到一定程度的阻碍,进而影响植物的生长发育和生理生化及其代谢[18]。本研究中4个杨树无性系不同干旱条件下各指标方差分析结果显示,除气孔器长在处理间差异不显著外,其余各性状在无性系间、处理间和无性系与处理交互作用间均达到极显著差异水平(P<0.01)。说明不同无性系间存在遗传差异,而且在不同干旱环境中其响应方式不同。依据不同的差异响应可对各参试杨树品种进行选择,以获得抗旱性优良的杨树材料。
光合作用是植物最基本的生命活动,其强弱是影响植物生长的主要因素[19],干旱胁迫下植物的净光合速率变化是植物对干旱胁迫适应能力的直接体现[20],叶片气孔导度的变化在一定程度上对植物的抗旱能力具有重要作用[21]。利用光合指标作为评价指标,对植物的耐旱能力进行鉴定,筛选出的材料具有较高的利用价值。本研究中,光合速率的降低表明干旱胁迫对植物光合系统造成一定程度的影响。部分研究表明干旱条件下植物光合速率降低,主要有两方面原因导致,一是气孔因素起主导作用,二是叶肉细胞中RUBP羧化酶活性降低导致[22~23]。气孔是植物与外界进行气体交换的门户,外界二氧化碳通过气孔进入植物的叶肉细胞,叶肉及表皮细胞的水分通过气孔蒸发到环境中[24]。植物蒸腾作用中气孔蒸腾占据重要地位,而气孔密度及大小与其蒸腾作用密切相关[25]。不同干旱胁迫处理下,除气孔器长差异不显著外,其余气孔形态指标均达极显著差异水平。各气孔指标均随土壤相对含水量的降低,呈现减小趋势。这与Tardieu和Tuberosa[26]对胁迫处理下气孔孔径和数量减少的结果相同。本研究中气孔形态指标间相关性达极显著水平与王锋堂等[27]对干旱胁迫下热带樱花的研究结果相同,表明植物通过改变气孔形态和密度来适应干旱的胁迫。另一方面,根据Farquhar等[28]观点,判断植物光合作用降低的原因是气孔或非气孔限制影响的因素是胞间二氧化碳浓度的变化趋势。本研究中导致光合速率变化的因素与付士磊等[29]研究结果相似,当土壤相对含水量大于40%时,4个杨树无性系胞间二氧化碳浓度随着水分的减少而持续降低,胞间二氧化碳浓度变化趋势与气孔导度和气孔限制值变化趋势一致,表明此时叶片光合速率下降是由叶片气孔部分关闭导致的;当土壤相对含水量低于40%时,出现上升趋势,说明此时叶片光合速率受非气孔限制因素影响。与此同时,土壤相对含水量的变化还会对植物的水分利用率产生重要影响。植物的水分利用率是植物利用水分的一个重要评价指标[30],是林木在干旱地区造林成活的主要因素之一。本研究中,苗木水分利用率随土壤相对含水量的逐渐降低大体呈现为先上升后下降的趋势,钟培芳[31]和陈超[32]等研究水分利用率变化趋势与其一致。植物水分利用率受蒸腾速率气孔等影响,本研究中,随着土壤相对含水量的降低,为维持基本代谢活动,4个杨树无性系蒸腾速率和气孔导度均随干旱胁迫程度的加重而呈现下降趋势,而水分利用率却呈现出不同变化,出现这种结果的主要原因可能是,气孔导度下降导致光合速率和蒸腾速率产生变化,叶片蒸腾速率下降速度较光合速率下降快所导致。部分研究表明高水分利用率是植物对水分亏缺的一种响应[33],也是植物应对干旱环境的一种表现[34],本研究中,当土壤相对含水量降至40%时,与对照相比白城小青黑杨和白林3号杨还具有较高的水分利用率,因此在水分匮乏环境下选用白城小青黑杨和白林3号杨水分利用率较高无性系具有重要意义。
植物在干旱胁迫下,会通过产生一系列变化来适应环境,对不同指标进行相关性分析表明,大部分指标间具有显著相关性,因此采用隶属函数法对不同处理下各无性系指标进行综合评价,综合分析表明,白城小青黑杨在各个干旱处理水平中均表现出相对较好的抗旱能力,可为东北干旱和半干旱地区的推广造林提供材料。综上所述,在东北地区水分条件较差的地区可以选择白城小青黑杨这个树种,以发挥其水分利用率高和综合抗旱能力强的特点来适应当地生境,而在水分较好的地区则可以发展白城小青黑杨、白城小黑杨和白林3号杨来获得最大效益。
