兰科新种巾唇兰的花粉团发育及分类学意义
2020-04-08李楚然王艳萍谭庆琴
李 璐 张 锦,2 李楚然 王艳萍,2 谭庆琴,2 罗 艳
(1.西南林业大学云南生物多样性研究院,昆明 650224; 2.西南林业大学生命科学学院,昆明 650224; 3.中国科学院西双版纳热带植物园园林园艺部,勐腊 666303)
自19世纪初以来,花药形态特征的多样性在兰科的系统演化中占有重要的一席之地。1810年Brown把兰科分成了五个组(secitons)时,就参考了花粉粒的愈合程度和排列方式[1]。到1826年,Lindley建立兰科的新属、新种、组和族等分类阶元时,花药特征成了主要分类依据[2]。花药发育特征的多样性一直是兰科分类鉴定和系统演化的重要依据,例如:成熟花药的散粉方式(花粉粒、粘性花粉粒、花粉小块、花粉团),花粉团数目(2,4,6或8)和形态(全缘、孔裂、深裂或浅裂)的形成机制,花药壁发育类型,小孢子发生和雄配子体发育的胚胎学特征等[3~6]。
巾唇兰属(PennilabiumJ.J.Sm.)是兰科(Orchidaceae)一类多年生附生兰,株型矮小,叶片扁平、肉质、丛生;总状花序侧生,由1~2朵花组成,花为白色、黄色、奶油色或橙色肉质,唇瓣与合蕊柱基部愈合,具距,三裂,中裂片较短肉质;侧裂片大,截形或退化为耳状,前端边缘具不规则的齿;具2个近圆形的花粉团[7~8]。巾唇兰属约有17种,全世界分布中心位于马来西亚半岛和婆罗洲[9~11]。巾唇兰属为中国分布的新记录属,研究资料较少。2004和2014年我国植物学家先后报道了巾唇兰属的2个种,即巾唇兰(P.yunnanenseS.C.Chen & Y.B.Luo)[7]和鸵鸟巾唇兰(P.struthioCarr)[11],前者为新种,发现于云南南部,在印度和泰国也有分布;后者为发现于海南的中国新记录,在马来西亚半岛、婆罗洲和越南也有分布[12]。
最近的分子系统学研究一致认为,巾唇兰属为兰科树兰亚科万代兰族指甲兰亚族(Subfam.Epidendroideae,Trib.Vandeae,Subtrib.Aeridinae)成员和我国也有分布的管唇兰属(Tuberolabium)、虾尾兰属(Parapteroceras)以及分布在热带亚洲的几个小属组成了一个稳定的单系类群,即囊唇兰近缘类群分支(Saccolabiumalliance)[13~17]。有意思的是,巾唇兰属建立时就是从囊唇兰属分离出来的,但后选模式种是由L.A.Garay指定为P.angraecum(Ridl.) J.J.Sm[18~19]。目前,关于巾唇兰属的研究资料比较少,需要更多的学科证据来澄清它和囊唇兰属的亲缘关系。
巾唇兰是附生于海拔约1 300米的茶树树干上的多年生小型草本,1987年在云南省景洪市发现后被移栽至西双版纳热带植物园,于2004年发表的一个巾唇兰属新种。正如O’Byrne所描述的那样,巾唇兰属所有的种都是罕见和稀有的[20],目前关于巾唇兰属的研究较少,亟待补充[7]。考虑到花粉团发育的胚胎学特征在系统进化、传粉生物学以及生殖生物学方面的重要性,本文通过花形态解剖和石蜡切片技术观察了巾唇兰的花形态特征和花粉团发育过程,为更好地理解巾唇兰属在指甲兰亚族中亲缘系统关系和人工繁育的保护生物学方面提供科学依据。
1 材料与方法
实验材料栽培于西南林业大学科研教学实习基地(云南丰春坊生物科技有限公司兰花资源圃),正常开花。在2018年7月采集不同发育时期的花蕾和开放花,一部分新鲜材料用于解剖镜观察拍照,另一部分固定在FAA保存。固定后的材料经系列酒精脱水、石蜡包埋、切片(切片厚度7 μm)、铁钒—苏木精染色、中性树脂封片,烘干后用NikonE600显微镜观察拍照。用图像编辑软件Adobe Photoshop CS6完成研究结果的图片排版。
2 结果与分析
2.1 花形态解剖特征
巾唇兰是多年生附生草本,茎直立,具3~4枚叶,扁平稍肉质长圆形,具宿存的鞘。总状花序侧生于茎的基部,具2~3朵花,花序近下垂(图版Ⅰ:1)。花瓣和花萼浅黄色,花瓣全缘、狭长圆形,先端渐尖,形状与萼片相似而稍小。唇瓣3裂(图版Ⅰ:2)。合蕊柱短,无蕊柱足(图版Ⅰ:3~6)。唇瓣贴生于蕊柱基部,无关节;侧裂片大,近匙形,前端边缘具不规则的齿(图版Ⅰ:3);肉质中裂片较短,近舌形,先端锐尖,中部有两条纵向褶皱带紫色斑点(图版Ⅰ:4~6),距圆筒形,末端圆形(图版Ⅰ:3~4)。在花药帽前端可见一个狭长2裂蕊喙(图版Ⅰ:5)。花粉块(pollinarium)由2个近球形的蜡质花粉团(pollinium)、粘盘柄和粘盘组成(图版Ⅰ:7~9)。粘盘质地较厚,椭圆形;粘盘柄从基部向上扩大成翼形(图版Ⅰ:9)。
2.2 巾唇兰的花粉团的发育
早期的花药原基分化为一对侧生并列的椭圆形花药室,并朝唇瓣方向弯曲生长,两个药室之间有明显的药隔组织相连,每个药室有一团染色较深的孢原细胞组成的孢子囊(图版Ⅱ:1~2)。每个孢子囊内,靠近两个药室之间的内侧造孢细胞的轮廓和细胞核比外侧的大(图版Ⅱ:1,3)。随花药的发育,孢子囊中央的孢原细胞分化为染色较浅的薄壁细胞,其细胞轮廓较大,染色浅,排列稀疏,发育为一团不育隔膜组织(图版Ⅱ:4~9)。孢子囊中央的不育隔膜组织并没有横贯花药室,仅与两个药室内侧的花药壁相连接(图版Ⅱ:10)。与此同时,不育隔膜组织周围的造孢细胞则发育为小孢子母细胞,继续花粉发育过程。在小孢子母细胞通过减数分裂形成小孢子四分体期间,随着花药壁中的绒毡层和中层细胞的降解,孢子囊和花药壁之间出现明显的空隙,同时不育隔膜组织也出现降解而在孢子囊中央形成空隙(图版Ⅱ:11)。此时,位于孢子囊中央的小孢子四分体的轮廓明显比外侧的细胞轮廓大(图版Ⅱ:12)。这样的细胞轮廓差异一直持续到2-细胞型花粉时期(图版Ⅱ:14~15)。雄配子体发育时期,同一个孢子囊内的小孢子四分体进行有丝分裂的过程是不同步的,靠外的小孢子四分体已经完成了有丝分裂形成了2-细胞型花粉,而内侧的依旧处于小孢子四分体阶段(图版Ⅱ:12,15)。在雄配子发育阶段,随着两个药室靠近的内花药壁的断裂消失,每个成熟的花粉团内的不育隔膜组织也被彻底吸收,形成了一对开口朝内的孔裂花粉团(图版Ⅱ:13~14)。
2.3 巾唇兰的花药壁发育
花药原基发育的早期,一层排列整齐的表皮细胞包围着一团染色较深的孢原细胞组织(图版Ⅲ:1)。随着花药的发育,表皮下的一个孢原细胞充当了初生周缘壁细胞的功能,通过平周分裂形成了两个切向排列的子细胞(图版Ⅲ:2),靠外的为外次生周缘壁细胞,靠内的为内次生周缘壁细胞(图版Ⅲ:3)。接着,外次生周缘壁细胞直接发育为药室内壁细胞,内次生周缘壁细胞进行第二次平周分裂,形成两个子细胞(图版Ⅲ:4)。在小孢子母细胞减数分裂前期,这两个子细胞进一步分化为中层和绒毡层(图版Ⅲ:5~6)。此时发育完整的花药壁,由外到内分别为表皮、药室内壁、中层和绒毡层,花药壁发育模式符合单子叶型[21]。此时,表皮细胞开始变得细窄,出现降解,药室内壁细胞方阔型,中层细胞细长,绒毡层细胞圆球形,单核。到小孢子母细胞减数分裂Ⅰ的末期,表皮细胞发生了变形和降解,药室内壁细胞出现了明显的带状纤维状加厚,中层和绒毡层细胞已经降解,花药壁和花粉团之间出现明显空隙(图版Ⅲ:7)。在小孢子四分体进行有丝分裂形成2-细胞型花粉阶段,花药壁只剩下纤维状加厚的药室内壁和逐渐降解消失的表皮细胞残迹(图版Ⅲ:8~9)。
2.4 巾唇兰的小孢子发育
早期花药原基的表皮细胞下包围着一团染色较深的分生组织细胞,经过多次有丝分裂形成相继分化为造孢细胞和孢原细胞。这两类细胞形态小、呈圆球形,排列紧密。接着,除了靠近两个花粉团之间的内侧花药壁的少量孢原细胞分化为不育隔膜细胞组织外,大部分孢原细胞直接发育为小孢子母细胞(图版Ⅱ:7)。小孢子母细胞呈多边形,细胞质浓厚,染色较深,逐渐液泡化,细胞壁外被明显的胼胝质(图版Ⅳ:1~3)。小孢子母细胞经过减数分裂第一次分裂后形成了没有子细胞壁的两个子核(图版Ⅳ:4),保持在同一个细胞壁内完成减数分裂Ⅱ(图版Ⅳ:5),产生了没有明显子细胞壁的4个子核,即小孢子四分体前体(图版Ⅳ:6~8)。随着发育的进行,四个子核之间出现明显的细胞壁,形成了排列多样的小孢子四分体(图版Ⅳ:9)。因此,巾唇兰的小孢子母细胞减数分裂的胞质分裂方式为同时型。小孢子四分体的排列多为正四面体、左右对称和线型,稀交叉型(图版Ⅳ:6~9)。在雄配子体发育阶段,小孢子四分体并没有散开,依旧处于同一个胼胝质内,通过有丝分裂形成了体积较大的营养细胞和体积较小的生殖细胞(图版Ⅳ:10)。随后,生殖细胞融入到营养细胞中,发育为2-细胞型或2-核的成熟花粉(图版Ⅳ:11)。成熟花粉以四合花粉的形式存在,两两紧密排列,构成了质地坚硬的花粉团(图版Ⅱ:13~14)。雄配子体发育过程中,部分花药室出现了花粉败育的现象,已形成的小孢子四分体未能继续有丝分裂,细胞轮廓模糊变形,发育停止(图版Ⅳ:12)。

图版Ⅰ 巾唇兰的花形态特征 1.总状花序;2.花形态正面图,示唇瓣3裂,中裂片短小,侧裂片长;3~6.移去花萼和花瓣后的合蕊柱(3)和仅保留了唇瓣中裂片的合蕊柱(4~6)的正面(5)和侧面(3,4,6),示合蕊柱短,花粉团一对,球形,具花药帽;唇瓣基部具圆柱形的花距(3~4),唇瓣侧裂片边缘具不规则的齿状(3),中裂片近舌型(4~5),唇瓣中裂片表面具纵向褶皱突起(白色箭头,4~6),花药帽前沿具一个二分叉的蕊喙结构(白色箭头,上侧,5);7~9.花粉团的背面(7)、侧面(8)和正面(9) ac.花药帽;c.合蕊柱;lip.唇瓣;lll.唇瓣侧裂片,mll.唇瓣中裂片;p.花粉团;sp.距;s.粘盘柄;v.粘盘 花粉团的这三个面皆全缘,无裂缝或孔。Plate Ⅰ Floral morphology in P.yunnanense 1.An inflorescence; 2.A flower in front view,showing a 3-lobed lip; 3-6.The column with sepals and petals removed(3),and with middle lobe of lip remained(4~6),in side views(3,4,6) and front view(3),showing a short colum,a cylindrical spur,two lateral lobes of lip with fimbriate margin(3),the middle lobe of lip(4,5) with longitudinal folds on upper surface(white arrows,4-6),the anther cap with a biforked beak at the front end(black arrow,5); 7-9. The back(7),side(8),and front(9) views of two pollinia,note that the pollinia are entire in these views,without cleft or pore ac.Anther cap; c.Column; lll.Lateral lobe of lip; mll.Mid lobe of lip; p.Pollinium; sp.Spur; s.Stipe; v.Viscidium
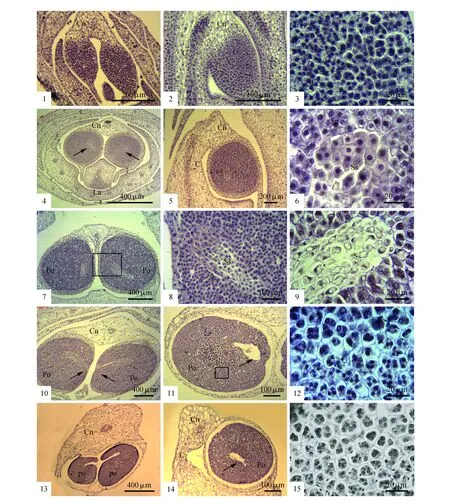
图版Ⅱ 巾唇兰的花粉团发育 1~2.早期的花药原基分化为一对侧生并列的椭圆形药室,朝唇瓣方向弯曲生长,一对花药室的横切图(1),单个花药室的侧面纵切(2);3.图1方框部分的放大图,示造胞细胞的轮廓在小孢子囊中央(右侧,3)的比外侧的大(左侧,3);4~10.小孢子囊中央的造孢细胞分化为一团染色较浅的薄壁组织细胞(箭头所示,4),细胞轮廓大,排列稀疏(6,8~9),发育为一段不横贯药室的不育隔膜组织(箭头所指,10);11~12.小孢子四分体时期,花药壁的中层和绒毡层降解消失,孢子囊和花药壁之间出现明显空隙,孢子囊内的不育隔膜组织也出现降解形成空隙(箭头所指,11),且小孢子囊中央的小孢子四分体(上部,12)轮廓明显比外围的大(下部,12);13~15. 2-细胞型花粉时期,形成两个开口朝内的孔裂花粉团(箭头所指,13),单个花粉团的侧面纵切图(14),此时同一个孢子囊内的花粉有丝分裂不同步,靠外的轮廓小的小孢子四分体已发育为2-细胞型四合花粉(底部,15),靠内的轮廓大的细胞尚处在小孢子四分体阶段(上部,15) Cn.药隔组织;C.合蕊柱;Po.花粉团;Sp.孢子囊;Ss.隔膜组织;La.唇瓣Plate Ⅱ Pollinia development in P.yunnanense 1-2. The anther primordium differentiated into a pair of lateral juxtaposed thecae bending toward to the lip,transverse section of a pair of thecae(1),and of a single theca in side view(2); 3. Magnified view of a part of fig.1,showing the sporogeneous cells in the center(the right side of fig.3) larger than those surrounding(the left side of fig.3); 4-10. The sporogenous cells in the center of the microsporangium differentiated into a group of light stained parenchyma(arrow,4),with large cells and arranged loosely(6,8,9),an incompletely sterile septum cross the thecae(arrow,10); 11-12. Microspore tetrad stage,the middle layer and tapetum degraded with an evident gap between sporangium and anther wall,the sterile septum degraded(arrow,11),noting that the microspore tetrads in the center of the microsporangium(the top of fig.12) is significantly larger than those surrounding(the bottom of fig.12); 13-15. The sterile septum completely disappeared,forming two pollinia with the stomium lying between two thecae(asterisk,13); longitudinal section of a single theca in side view(14),asynchronism in male gametogenesis,noting that the tetrads surrounding at 2-celled pollen stage(the lower side of fig.15),while those in the center microspore at the microspore tetrad stage(the top of fig.15) Aw.Anther wall; Cn.Connective tissue; C.Column; Po.Pollinium; Ss.Sterile septum; St.Sporogenous tissue; La.labellum

图版Ⅲ 巾唇兰的花药壁发育 1.早期的花药原基为一层排列整齐的表皮细胞包围着一团染色较深的孢原细胞细胞组织,注意表皮细胞正进行垂周分裂(箭头);2~3.表皮细胞下的一个孢原细胞正在平周分裂产生两个子细胞(箭头,2),靠外的为外次生周缘壁细胞,靠内的为内次生周缘壁细胞(3);4.小孢子母细胞时期,外次生周缘壁细胞发育为药室内壁,内次生周缘细胞通过一次平周分裂,形成两个子细胞(箭头);5~6.减数分裂前期,花药壁发育完整,由外到内分别为表皮、药室内壁、中层和绒毡层。7.减数分裂后期,表皮、中层和绒毡层已经降解,药室内壁增大且发生纤维状加厚;8~9.小孢子四分体(8)和2-细胞花粉时期的花药壁,表皮细胞逐渐降解,药室内壁继续保持不均匀纤维状加厚 Ep.表皮;En.药室内壁;Isp.内次生周缘细胞;Ml.中层;Osp.外次生周缘细胞;Ta.绒毡层Plate Ⅲ The development of anther wall in P.yunnanense 1. A densely-stained sporogenous tissue surrounded by a layer of epidermal cells,noting that the epidermal cells were undergoing anticlinal division(arrows); 2-3. The hypodermal archesporial cells undergo a periclinal division resulting two daughter cells(arrow in 2),including the outer and the innner secondary parietal cells; 4. At the stage of microsporocyte,the outer secondary parietal cells function as endothecium,while the inner ones keep on producing two daughter cells(arrow); 5-6. The completed anther wall consisting of 4 layers,including epidermis,endothecium,middle layer,and a tapetum; 7. At the end of meiosis Ⅱ,the middle layer and tapetum were degraded,fibrous thickening occurred in the endothecium; 8-9. Anther wall at microspore tetrad stage(8) and 2-celled pollen stage,noting the epidermis gradually degraded,and endothecium became fibrous thickened Ep.Epidermis; En.Endothecium; Isp.Inner secondary parietal layers; Ml.Middle layer; Osp.Outer secondary parietal layer; Ta.tapetum
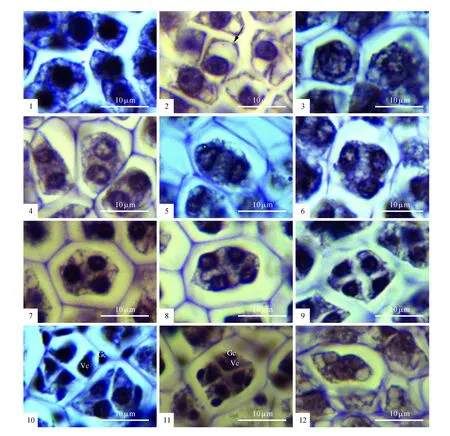
图版Ⅳ 巾唇兰的小孢子发育 1.孢原细胞;2.小孢子母细胞,细胞质内出现明显的液泡(箭头);3~9.小孢子母细胞减数分裂过程:减数分裂I前期(3),减数分裂Ⅰ后期(4)和减数分裂Ⅱ前期,注意未形成子细胞壁(5),减数分裂Ⅱ后期(6),减数分裂Ⅱ末期,小孢子四分体尚未形成细胞壁,4个子核排列呈正四面体(7)或左右对称(8);形成子细胞壁的小孢子四分体呈左右对称排列(9);10~11.雄配子体发育过程,小孢子四分体保持在同一个胼胝内继续有丝分裂,形成小的生殖细胞和大的营养细胞(10),接着,生殖细胞与营养细胞之间的细胞壁溶解后发育为2细胞型花粉(12);12.部分花药内靠近外侧花药壁的小孢子四分体未能继续完成有丝分裂,而出现败育 Gc.生殖细胞;Vc.营养细胞Plate Ⅳ The Microsprogenesis in P.yunnanense 1.Sporogenous cells; 2.Microspore mother cells; 3-9.Meiosis of microspore mother cells: Prophase Ⅰ(3),Anaphase Ⅰ(4),Prophase Ⅱ(5), Anaphase Ⅱ(6-8),noting the arrangements of microspore tetrads without cell walls in tetrahedral(6,7),isobilateral(8); 9.Microspore tetrads arranged in isobilateral type,noting the cell walls formed; 10~11.The microspore underwent a mitotic division and developed into 2-celled pollen; 12.Abortion of pollen at the stage of microspore,noting the distorted cells Gc.Generative cell; Vc.Vegetative cell
3 讨论
本文通过解剖镜和石蜡切片技术,观察了巾唇兰的花形态和花药发育过程,从成熟花的形态解剖特征和花药发育的组织切片水平揭示了一些具有重要分类学意义的胚胎学特征,具体如下。
第一,本研究从组织切片水平揭示了巾唇兰两个孔裂花粉团的形成机制,证实了早期花药发育过程中,小孢子囊内孢原细胞组织分化出不同类型的不育隔膜组织对兰科中花粉团的数目和形态特征的形成具有决定性作用[22]。巾唇兰的隔膜组织的发育模式很特别,隔膜组织起源于药室中央分化为不育隔膜组织的一团孢原细胞。随着花药生长,这段隔膜组织一端始于小孢子囊中央,另一端延伸至两个药室相邻的内侧花药壁。在发育过程中,该隔膜组织逐步降解,至花药成熟时彻底消失,在两个药室相邻的内侧花药壁断裂处形成孔隙,发育为一对开口朝内的孔裂花粉团。已有研究表明,在兰科不同的类群中,隔膜组织分化的有无、数目及发育朝向对于花粉团的不同数目及其表面形态特征的形成机制具有重要作用[22~23]。在树兰亚科的花粉团发育过程中,如果花药室内没有出现隔膜组织,就会形成2个实心花粉团,如Stelisciliaris;如果花药室内发生的部分分隔(没有形成完整的隔膜组织),可能形成浅裂花粉团(lobed pollinia),如合萼兰属(Acriopsis)或具孔的花粉团;如果每个花药室内都形成一个背腹向隔膜组织,将形成最常见的4个花粉团,如沼兰属(Malaxis),石仙桃属(Pholidota),盖喉兰属(Smitinandia);而虾脊兰属(Calanthe)等8个花粉团可由两条纵向或一纵一横的隔膜组织切割而成[22~23]。
第二,从个体发育组织解剖水平,本文澄清了巾唇兰的花粉团具孔裂,而非如前人所描述的那样全缘[8,13~14]或实心[10]。在解剖镜下,巾唇兰成熟花药中有一对由粘盘和粘盘柄相连接的球形花粉团,该花粉团从正面、侧面和背面皆为全缘,无孔裂或浅裂,这与前人的描述是一致的。然而,本文通过石蜡切片观察到成熟花药的纵切面上,每个药室内的花粉团在靠近两个药室相邻的内花药壁处,存在着一条开口朝内花药壁处的空隙。这说明巾唇兰的花粉团并非全缘或实心,而是孔裂。花粉团孔裂的特征,从花药发育过程的组织切片上可以得到追踪和证实。早期花药原基分化出的一对侧生药室内,其中央分化出一段不育隔膜组织,始于药室中央,止于两个药室相邻的内侧花药壁组织。在花药成熟时,两个药室相邻的内侧花药壁发生降解形成药室裂口,同时这段不育隔膜组织也被全部吸收,在两个成熟的花药室相邻的地方形成一道开口朝内的一对孔裂花粉团。由于花粉团的孔裂开口位于两个花粉团紧密相连处,在解剖镜下不易观察到,但在组织切片水平上却可以完整地看到该孔裂的形成机制。因此,从这个角度而言,对于具有不同形态特征的花粉团来说,一方面需要全方位仔细观察成熟花粉团的形态特征,另一方面对于开口较小或不易察觉的孔裂或浅裂的花粉团,可以通过组织切片技术来进一步澄清。
第三,巾唇兰两个孔裂花粉团的形成机制,对于万代兰族2个花粉团特征及其形成机制具有重要的启示作用。在巾唇兰所处的万代兰族指甲兰亚族的类群中,花粉团的数目和形态特征描述在近缘类群甚至一个属内都不一致,无论是传统的形态分类和分子系统学都很难用来反映它们的系统亲缘关系[13~15,24]。根据前人的研究,一些在分子系统树上亲缘关系比较近的指甲兰亚族类群,往往具有不同数目和形态特征的花粉团的描述[13~15,24]。例如,在巾唇兰属所在的囊唇兰近缘类群分支中,巾唇兰同我国分布的囊唇兰和管唇兰以及其他2个小属(Ceratocentron,Hymernorchis)都被描述为具有一对全缘的花粉团,但在其它属里却分别具有孔裂花粉团孔裂花粉团(如:Dyakia,Cryptophylos)和浅裂花粉团(如:Macropodanthus,Amesiella)。在指甲兰属的近缘类群中,风兰属(Neofinetia)和槽舌兰(Holocoglossum)具有一对孔裂花粉团;但万代兰属和钻喙兰属(Rhyncostylis)以及国外的一个小属(Trudelia)具有一对带裂缝的花粉团。甚至在同一个属里花粉团也被描述为既有孔裂和浅裂,例如指甲兰属和鸟舌兰属。除了在同一个近缘类群分支或同一个属中,花粉团的形态特征有不同的描述之外,花粉团的数目也有变异,例如:Pomatocalpa的近缘分支类群中,Pomatocalpa有4个,Haraella有2个;同样,Diploprora的近缘类群中,Diploprora有4个花粉团,而另外两个属(Omoea,Malleola)有2个。在蝴蝶兰属中,花粉团数目也有2个或4个的描述。面对万代兰指甲兰亚族的花粉团形态特征的不同描述给分类系统带来的困惑[13~15,24],本研究结果说明对于万代兰族指甲兰亚族错综复杂的分类系统学问题,对花粉团的不同的数目或形态特征形成机制,有必要从花药发育的个体形态解剖水平上进一步研究,准确判断花粉团的形态类型。
第四,巾唇兰的花距、无蕊柱足、花粉团形态、粘盘等特征在属间及种间具有重要的分类学意义。巾唇兰属是J.J.Sm.于1914年由囊唇兰属中分隔出来的[18~19],通过对巾唇兰花形态解剖特征观察,支持J.J.Sm.将巾唇兰属由囊唇兰属中分隔出来。从花形态解剖特征来看,巾唇兰和囊唇兰的区别显著,巾唇兰的花距明显,为长圆柱形,无蕊柱足,具有一对孔裂花粉团;而囊唇兰的唇瓣基部的花距短,为圆筒状距形,有蕊柱足,花粉团为全缘[25~26]。因此,花距长圆柱形、花粉团孔裂,蕊柱足不明显等形态解剖特征支持巾唇兰属应该从囊唇兰属中分离出来。此外,根据本文的研究结果,我国产的两种巾唇兰属植物在花形态特征方面较为相似[11],球形花粉团,椭圆形的粘盘以及由上至下扩大呈翼型或长三角型条带的粘盘柄,明显的区别在于巾唇兰的花梗和子房旋转,而鸵鸟巾唇兰的不旋转[11]。显然,关于和巾唇兰属一样曾被描述为具有2个全缘花粉团的近缘属(管唇兰属,火炬兰属,虾尾兰属)来说,花粉团是否全缘或孔裂这一关键分类学特征也需要花粉团发育的解剖证据来进一步澄清。
第五,巾唇兰花药发育过程中的一些胚胎学特征反映了兰科花药发育的多样性和普遍性,为更好地理解兰科分类系统和演化提供新证据。例如,单子叶型的花药壁发育类型、表皮细胞的过早降解、单核绒毡层、同时型胞质分裂、小孢子四分体排列形式、雄配子体发育过程中的同步性以及2-细胞型的成熟花粉等特征在不同的亚科、族和属间都明显不同,具有一定的分类学意义。
首先,巾唇兰的单子叶型花药壁发育方式和表皮细胞不宿存等特征丰富了兰科花药发育的多样性。巾唇兰成熟的花药壁有4层细胞,发育方式符合单子叶型。目前,兰科植物花药壁发育类型有两种:单子叶型和多层型(massive type)。单子叶型花药壁是指发育完整的花药壁有4层细胞组成:表皮、药室内壁、中层和绒毡层[21]。单子叶型花药壁在树兰亚科的万代兰族见于万代兰属(Vanda)、指甲兰属(Aerides)、钻喙兰属[27],在其他类群中还见于小斑叶兰(Goodyerarepens)[28]、沼兰属(Malaxisversicolor、M.muscifera)[29~30]、Microstyliscylindrostachya[31]、线柱兰属的Zeuxinestrateumatica[32]等。多层型花药壁是在羊耳蒜属(Liparis)植物中首次提出来的,其发育完整的花药壁有5~8层,远远超过了被子植物中其他类型(基本型,双子叶型,单子叶型,退化型)花药壁的组成层数,因此被命名为新的花药壁类型:多层型[33]。多层型花药壁层数的增加源于药室内壁或中层层数的增加,可多达2~5层。因此,多层型花药壁层数可达6~10层以上。多层型花药壁还见于杓兰亚科的杓兰属(Cypripedium)[34],兰亚科的绶草属(Spiranthus)和兰属(Cymbidium)[35]以及树兰亚科的苞舌兰属(Spathoglottis)[36]。因此,花药壁发育类型在兰科分类系统中具有重要作用。同时,巾唇兰的花药壁的表皮细胞并非宿存,表现为过早降解。这个发育特征在兰科植物中比较少见。表皮细胞在小孢子母细胞进行减数分裂时,表现为被拉伸和压缩,细胞轮廓变得细长,出现降解的迹象,在花药成熟时,花药壁仅剩药室内壁和部分表皮的残迹。这意味着巾唇兰的表皮细胞过早降解。然而,在现有的兰科花药发育的资料中,表皮细胞通常宿存[37~39]。关于表皮细胞的过早退化或降解的分类学意义,有待于更多资料的补充。
其次,巾唇兰花药发育过程中的一些特征与大部分兰科的相似,反映了兰科花药发育的普遍性。例如,巾唇兰小孢子母细胞减数分裂的胞质分裂方式为同时型,在兰科资料中较为常见。现有资料表明,兰科花药发育过程中的减数分裂的胞质分裂有两种:同时型和连续型。前者较为普遍,在早期大部分兰科植物都很普遍[40];后者较为少见,见于陆续报道的部分兰科,如:树兰亚科的虎舌兰属(Epipogium)[41]和苞舌兰属[36],兰亚科的眉兰属(Ophrys)[42]和绶草属[43]。大部分兰科植物中的绒毡层为腺质型,且普遍为单核,这在巾唇兰中也不例外。然而,在部分兰科成员中,也有双核绒毡层,如杓兰亚科的兜兰属(Paphiopedilumdruryi)[27]、树兰亚科的火烧兰属(Epipactislatifolia,E.veratrifolia)[44]。巾唇兰的小孢子四分体排列形式多样,多为正四面体和左右对称,稀十字交叉型或线型,且成熟花粉粒为2细胞型。以上这些特征在现有兰科资料中都较为常见[40],可以算作兰科花药发育的普遍特征。
此外,在巾唇兰花药发育过程中,同一个药室内存在着花粉败育现象。败育时间主要发生在小孢子四分体时期,靠近花药壁的小孢子四分体细胞轮廓变形,细胞质细胞核不清晰,出现败育现象,而处于药室中央的小孢子四分体形态正常,继续发育。这样的败育现象也许因为花药壁的绒毡层和中层细胞已经完全降解,因此靠近花药壁的小孢子四分体无法继续获得更多营养继续发育而出现败育。相反,靠近药室中央的孢子囊,由于隔膜组织细胞层数多,尚未降解完全,依旧可以继续为小孢子四分体进一步发育为雄配子体而提供营养,因此发育正常。同时,考虑到兰科花药内大量的花粉聚集成团,数目多达上百万,由于营养供给有限,导致了部分花粉的败育。由于花药发育过程中,存在花粉败育现象,导致了并非所有的小孢子母细胞都可以顺利发育为成熟花粉,限制了兰科双受精过程所必需的雄配子体数量的供给。因此,巾唇兰花粉团发育过程中存在的花粉败育现象,也从一定角度解释了野生兰科植物种群数量稀少,除了人为活动干扰外,还存在着一定的生殖机制障碍。
综上所述,本文首次为新种巾唇兰的花粉团发育提供了重要的胚胎学特征,为兰科植物的系统演化和生殖生物学提供了科学依据。然而考虑到兰科多达800属26 000种的庞大成员,且在亚科内的不同族、亚族和属间的系统亲缘关系亟待澄清[16~17],因此选择具有代表性的兰花观察花药发育特征,进行比较胚胎学研究是多多益善的[27~28]。
致谢本研究依托西南林业大学云南生物多样性研究院提供实验平台完成;西南林业大学科研教学实习基地(云南省丰春坊生物科技有限公司兰花基地)提供实验材料;西南林业大学古旭老师在实验过程中提供帮助。
