水稻条纹花叶病毒对介体电光叶蝉生长繁殖及取食行为的影响
2020-03-31陈红燕魏太云贾东升
李 盼, 张 洁, 岳 玥, 陈红燕, 吴 维, 魏太云, 贾东升,*
(1. 福建农林大学植物保护学院, 福建省植物病毒学重点实验室, 福州350002;2. 云南省农业科学院生物技术与种质资源研究所, 昆明 650000)
植物病毒是农业生产中的重要病原物,全球目前已报道的植物病毒约1 100种,其中约80%是由特定的介体昆虫传播,由于缺乏有效的防治方法,病毒病给农业生产造成了重大的损失(Ng and Falk 2006; Hogenhoutetal., 2008; 叶健等, 2018)。在农作物病毒病害的流行发生中,介体昆虫发挥着至关重要的作用,且植物病毒-介体昆虫-寄主植物三者间形成了复杂的互作关系。首先,由于病毒与介体昆虫的多样性,植物病毒对介体昆虫既存在有利的,也存在不利的影响(Hogenhoutetal., 2008; Whitfieldetal., 2015)。如感染水稻矮缩病毒(rice dwarf virus, RDV)的水稻提高了其介体黑尾叶蝉Nephotettixcincticeps的存活率和产卵量,促进种群增长(王前进等, 2018);而南方水稻黑条矮缩病毒(southern rice black streaked dwarf virus, SRBSDV)会显著降低介体白背飞虱Sogatellafurcifera若虫的存活率、延长若虫的发育历期及降低成虫寿命和繁殖力(Tuetal., 2013)。其次,植物病毒可以通过影响介体昆虫的取食行为而促进其自身的传播 (Fereres and Moreno 2009; Dietzgenetal., 2016; Shikanoetal., 2017),如植物病毒通过影响寄主植物的代谢过程或改变其性状,直接或间接使介体昆虫对寄主植物的吸引、选择、定居和取食等行为发生变化,进而促进病毒在寄主植物间的传播以及病害的远距离扩散(Moreno-Delafuenteetal., 2013; Luetal., 2016)。因此,植物病毒与介体昆虫之间的互作关系成为影响田间病毒病流行的重要因素(Stoutetal., 2006)。
水稻条纹花叶病(rice stripe mosaic disease)是2015年在我国广东省罗定市被首次发现的水稻新病害,感病水稻出现植株矮化、叶片条纹状花叶、新叶扭曲,结穗率下降等症状(Yangetal., 2017a)。该病害由水稻条纹花叶病毒(rice stripe mosaic virus, RSMV)引起,RSMV属于弹状病毒科(Rhabdoviridae)细胞质弹状病毒属Cytorhabdovirus的一个新种,主要由半翅目的电光叶蝉Reciliadorsalis以持久增殖型方式传播(Yangetal., 2017b),二点黑尾叶蝉Nephotettixvirescens是其次要传播介体(Zhaoetal., 2019)。目前该病害已扩散到广西、海南和云南等省,由于电光叶蝉种群繁殖能力强,扩散速度快,使该病害存在进一步蔓延的趋势(Yangetal., 2018)。近年来的研究已明确RSMV的基因组特征,RSMV由介体电光叶蝉传播的特性,以及RSMV在介体昆虫体内的侵染过程(Yangetal., 2017a, 2017b; Zhaoetal., 2019),而关于RSMV如何影响电光叶蝉的生长繁殖和取食行为尚不清楚。
为此,本研究将通过生物学实验测评RSMV对电光叶蝉生长发育的影响;利用EPG技术比较感染和未感染RSMV的电光叶蝉在水稻上的取食行为,探究RSMV对电光叶蝉取食行为的影响;采用Y型嗅觉仪研究电光叶蝉对感染和未感染RSMV的水稻的偏好性差异。从而明确电光叶蝉-寄主植物-病毒三者之间的相互关系,揭示RSMV扩散流行的机制,为建立有效的病毒病害防控技术体系奠定理论基础。
1 材料与方法
1.1 供试昆虫和水稻
电光叶蝉采自广东省罗定市,经实验室筛选繁殖后,无毒电光叶蝉在人工气候室(温度26±1℃,光周期16L∶8D,相对湿度60%~70%)中饲养于健康的感虫水稻品种TN1上,用于电光叶蝉寄主选择行为和生长发育实验。
感染RSMV的水稻采自广东省罗定市,种植于实验室水稻病毒圃。电光叶蝉2龄若虫在感染RSMV的水稻病株上取食2 d后转移至TN1水稻幼苗上饲养,获得电光叶蝉获毒种群,获毒后10 d的电光叶蝉用于EPG实验。同时将获毒后10 d的电光叶蝉向三叶期的日本晴水稻幼苗进行传毒,传毒后30 d通过症状观察并参照Yang等(2017b)RT-PCR方法检测水稻带毒情况,鉴定后的带毒水稻供试。同时以未进行传毒的日本晴水稻作为健康水稻供试。
1.2 RSMV侵染后电光叶蝉生长和生命表参数的测定
为了明确RSMV对电光叶蝉生长的影响,取RSMV侵染的水稻病株放入饲毒笼中,接入孵化不足12 h的电光叶蝉若虫约200头取食12 h。同时用棉花包裹10 d苗龄的稻苗根部并装入大试管中(长25 cm,直径3 cm),每试管1 株水稻,试管底部加适量水。接入毒株上取食12 h的若虫1头,以带纱网的试管盖封口。每日同一时间更换水稻幼苗1次,每12 h观察和记录若虫的蜕皮情况,直至羽化为成虫(Chenetal., 2016),并计算出不同龄期的发育历期。收集羽化后的成虫,并参照Yang等(2017b)RT-PCR方法检测电光叶蝉带毒情况,筛选出100头带毒电光叶蝉。同时以在健康水稻上取食后的100头若虫作为对照组进行实验。电光叶蝉饲养于恒温培养箱中,培养条件为光周期16L∶8D、相对湿度60%~70%、温度26±1℃。实验重复3次。
为了明确RSMV对电光叶蝉种群繁殖的影响,按以上方法取400头刚孵化的电光叶蝉若虫至装有1株10 d苗龄稻苗的大试管中单头饲养,每日同一时间更换水稻幼苗1次。逐日观察,并记录1-5龄若虫的存活数,以及羽化出的成虫和雌虫数。将羽化的成虫雌雄配对放入一个装有1株10 d苗龄稻苗的大试管中,每天更换稻苗,更换的水稻苗置于解剖镜下解剖,记录卵粒数,并将产的卵挑选到滤纸上孵化,统计孵化的卵数。在此过程中,若有雄虫死亡,及时补充新的电光叶蝉雄虫,直至雌虫全部死亡。没有产卵或所产卵均不能发育的雌虫视为交配失败。收集死后的成虫并参照Yang等(2017b)RT-PCR方法检测带毒情况,筛选出带毒电光叶蝉数据,计算存活率Su1-5、羽化率(Emr)、雌性率(Fr)、交配率(Cr)、繁殖力(Fy)和卵孵化率(Ehr)。根据种群起始数量(No),预测下代种群数量(Nt)和种群趋势指数(I),计算公式为:Nt=No×Su1-5×Emr×Fr×Cr×Fy×Ehr;I=Nt/No。随机取100头健康的电光叶蝉1龄若虫作为对照组进行实验。实验重复3次。
1.3 刺吸电位(EPG)记录与分析
利用DC-EPG Giga-8(荷兰瓦赫宁根大学)刺吸电位技术监测感染和未感染RSMV电光叶蝉口针在健康水稻上的取食行为差异。供试电光叶蝉成虫饥饿处理2 h后,在CO2短暂麻醉条件下迅速将一段2~3 cm、直径10 μm的金丝一端粘连在电光叶蝉的前胸背板上,另一端与EPG工作传感器的铜线粘连,同时植物电极插在水稻生长的土壤中。当电光叶蝉口针刺入水稻组织时,回路接通,电流信号经电流放大器放大后保存于电脑上,再用Probe 3.4软件转化为波形图谱输出在电脑屏幕上保存、判断并转换为可供统计的数字文件(万贵钧, 2015)。每头成虫记录5 h,每次记录结束后更换昆虫和水稻植株,感染RSMV和无毒电光叶蝉的处理组各测定15头成虫。实验在25±1℃,相对湿度60%±10%的安静房间内进行。
电光叶蝉取食波形判定参照陈勇(2015)关于电光叶蝉取食波形的分析进行鉴定,EPG的各种波形代表刺吸式传毒昆虫的口针在寄主植物组织内不同的刺探和取食行为,具有不同的生物学意义:NP波为非刺探波形,表示电光叶蝉停留在叶片上没有刺探;A波代表试探性刺探,一般持续时间比较短;C波代表口针刺探到韧皮部之前的主动取食波;E波表示昆虫口针在韧皮部内的被动取食;F波表示昆虫在刺探时的机械穿刺阻碍波;S波是刺探波之后的唾液分泌波;R波是休息波。波形分析采用Stylet软件,记录并输出每一个波形从开始到结束的时间,并进行数据分析。
1.4 电光叶蝉寄主选择行为的生物测定
使用Y型嗅觉仪测定电光叶蝉成虫对水稻的取食偏好性。取处于分蘖期的健康水稻和感染RSMV的水稻,分别放入2个味源瓶中,控制装置流量为1.0 L/min。将30头健康电光叶蝉成虫引入Y型管的管柄中,待其爬至两臂夹角处开始计时,观察6 h内电光叶蝉对两臂味源瓶的选择趋向(Wangetal., 2014)。6 h后停止行为观察,记录爬至各味源瓶中的电光叶蝉头数。同时,取30头饲毒后10 d的电光叶蝉成虫引入Y型管的管柄中,按以上方法测试带毒电光叶蝉对不同水稻的取食偏好性。6 h后统计爬至各味源瓶中的电光叶蝉数目,并收集虫体,参照Yang等(2017b)RT-PCR方法检测带毒虫数。每次测定更换Y型管,调换味源瓶方向,消除误差,每次试验重复3次。
1.5 数据分析
本研究所有数据均使用GraphPad Prism 7.0进行数据处理及分析。采用T检验比较分析电光叶蝉在健康与感病水稻上的发育历期、成虫寿命、羽化率及产卵量等。同时采用T检验比较分析电光叶蝉对健康与感病水稻的选择偏好性以及RSMV对电光叶蝉成虫取食波形出现次数和持续时间的影响。
2 结果
2.1 RSMV对电光叶蝉生长繁殖的影响
对不同龄期电光叶蝉若虫在健康水稻幼苗上的发育历期进行统计,获得感染RSMV电光叶蝉若虫和无毒电光叶蝉若虫的1, 2, 3, 4和5龄若虫期以及总若虫期(表1)。其中带毒电光叶蝉1, 2, 3和5龄以及若虫总发育历期均较无毒电光叶蝉若虫发育历期显著延长(P<0.05),仅4龄若虫发育历期无显著差异(P>0.05)(表1)。表明RSMV侵染电光叶蝉后显著延长了电光叶蝉若虫的发育历期。
系统分析RSMV对电光叶蝉生命表的影响,发现带毒电光叶蝉的若虫存活率为65.75%,显著低于无毒电光叶蝉的79.22%;带毒电光叶蝉的羽化率为84.02%,显著低于无毒电光叶蝉的90.56%(P<0.05);带毒电光叶蝉的繁殖力为58.38粒/雌虫,显著低于无毒电光叶蝉的85.44粒/雌虫(P<0.05);带毒电光叶蝉的卵孵化率为60.28%,显著低于无毒电光叶蝉的67.55%(P<0.05);此外,带毒电光叶蝉的雌性率和交配率与无毒电光叶蝉没有显著差异(P>0.05)。综合计算带毒电光叶蝉的F1代种群趋势指数为8.45,显著低于无毒电光叶蝉的19.48(P<0.05)(表2)。以上结果表明RSMV侵染电光叶蝉后显著降低了昆虫的若虫存活率、羽化率、繁殖力和卵孵化率,从而显著降低了F1代的种群趋势指数,表明RSMV不利于电光叶蝉种群的繁衍。

表1 水稻条纹花叶病毒侵染对电光叶蝉若虫发育历期的影响Table 1 Effects of rice stripe mosaic virus (RSMV) infection on the nymphal duration of Recilia dorsalis

表2 水稻条纹花叶病毒侵染对电光叶蝉生命表参数的影响Table 2 Effects of rice stripe mosaic virus (RSMV) infection on the life table parameters of Recilia dorsalis
2.2 RSMV感染对电光叶蝉成虫取食过程各波形出现次数和持续时间的影响
对带毒和无毒电光叶蝉进行刺吸电位波产生次数和持续时间进行分析和统计,发现携带RSMV的电光叶蝉比无毒电光叶蝉取食过程中产生A, F, S和NP波的次数显著增加(P<0.05),产生C和E波的总次数没有明显差异(P>0.05),而产生R波的次数显著下降(P<0.05)(图1: A)。同时对不同取食波形持续时间进行统计,发现携带RSMV的电光叶蝉产生A, C, F, S, R和NP波的时间显著高于无毒电光叶蝉(P<0.05),而产生的E波总时间显著低于无毒电光叶蝉(P<0.05)(图1: B)。综合不同波形产生的次数和持续时间,以及波形所代表的取食行为,表明携带RSMV的电光叶蝉在水稻上刺探取食时增加了障碍波和刺探波的次数,以及唾液分泌波的次数和时间;同时电光叶蝉被动取食行为减少,休息波次数减少,但每次休息时间延长。
2.3 RSMV对电光叶蝉成虫寄主选择行为的影响
通过Y型嗅觉仪对电光叶蝉成虫的寄主选择行为进行测定,在无毒电光叶蝉成虫中,约11头选择RSMV侵染的水稻,约5头选择健康水稻,表明无毒电光叶蝉成虫极显著趋向于选择RSMV侵染的水稻(P<0.01)。同时在获取RSMV的电光叶蝉成虫中,经检测约8头选择健康水稻,约2头选择RSMV侵染的水稻,表明带毒电光叶蝉极显著趋向于选择健康水稻(P<0.01)(图2)。
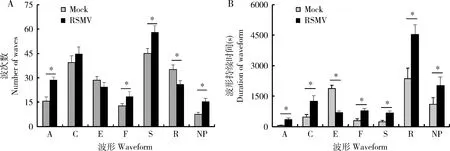
图1 水稻条纹花叶病毒对电光叶蝉成虫取食波形出现次数(A)和持续时间(B)的影响Fig. 1 Effects of rice stripe mosaic virus (RSMV) infection on the number of waves (A) and duration (B) of each feeding waveform for Recilia dorsalis adultsA: 刺探波Probing wave; C: 主动取食波Active ingestion wave; E: 被动取食波Passive ingestion wave; F: 障碍波Obstructive wave; S: 唾液分泌波Saliva excreting wave; R: 休息波Resting wave; NP: 非刺探波Non-probing wave; Mock: 无毒电光叶蝉Nonviruliferous R. dorsalis; RSMV: 带毒(RSMV侵染)电光叶蝉Viruliferous (RSMV-infected) R. dorsalis. 图中数据以平均值±标准误表示;星号表示电光叶蝉在健康水稻和RSMV侵染水稻上产生不同取食波形出现次数或持续时间差异显著(P<0.05, T检验)。Data in the figure are mean±SE. The asterisk indicates significant difference in the number of waves and duration of each feeding waveform between R. dorsalis adults feeding on healthy and RSMV-infected rice plants (P<0.05, T-test).

图2 携带 RSMV和无毒电光叶蝉成虫对健康和感病水稻的选择Fig. 2 Host plant selection of RSMV-infected and non-infected Recilia dorsalis adults to healthy and RSMV-infected rice plants图中数据以平均值±标准误表示;双星号表示电光叶蝉成虫在健康水稻和RSMV侵染水稻间选择性取食的差异极显著(P<0.01, T检验)。Data in the figure are mean±SE. The double asterisk indicates extremely significant difference in the selection of R. dorsalis adults to healthy and RSMV-infected rice plants (P<0.01, T-test).
3 讨论
植物病毒的传播是基于病毒-宿主-介体间三者互作关系的复杂过程(Dáderetal., 2017)。植物病毒对介体昆虫的影响直接关系到病害的扩散和蔓延(Maucketal., 2012)。本研究结果发现RSMV侵染延长了电光叶蝉若虫的总发育历期。基于电光叶蝉若虫的获毒和传毒效率均高于成虫(Yangetal., 2017b),推测病毒侵染延长若虫的发育历期对病毒的传播是有利的。同时发现RSMV对电光叶蝉的存活率、羽化率和繁殖力有显著影响,与无毒电光叶蝉相比,RSMV侵染的电光叶蝉存活率下降,羽化率降低,产卵率显著下降,导致种群趋势指数显著下降。这与水稻瘤矮病毒(rice gall dwarf virus, RGDV)对电光叶蝉生长繁殖的影响(Chenetal., 2016)相似,而不同于RDV有利于黑尾叶蝉种群的繁殖(王前进等, 2018)。表明电光叶蝉作为主要介体传播RSMV和RGDV的过程中,病毒的侵染降低了电光叶蝉的生长繁殖能力而不利于其种群的增长。
以持久增殖型方式传播的植物病毒需在昆虫体内增殖并扩散到唾液腺,随着唾液的分泌传播到健康植物(Hogenhoutetal., 2008)。植物病毒在昆虫体内的增殖对介体昆虫的取食行为产生直接的影响。本研究通过EPG测定发现RSMV感染的电光叶蝉在健康水稻上刺探取食的刺探波和取食时唾液分泌波次数显著增多,同时在水稻上刺探取食产生的障碍波次数和时间也增多,表明病毒侵染降低了电光叶蝉在植物韧皮部成功取食的能力,需要更多的刺探尝试。这些因素的变化有利于介体电光叶蝉在刺探过程中将携带病毒的唾液分泌到植物中,从而提高病毒的传播效率(施艳等, 2013)。该现象与RGDV对电光叶蝉取食行为的影响(陈勇, 2015)相同,推测一方面病毒侵染的电光叶蝉可能需要获取更多的能量物质维持除自身需要外病毒增殖的需求,另一方面病毒侵染降低了电光叶蝉成功取食的能力,产生更多的障碍波,这也可能是病毒侵染导致电光叶蝉种群繁殖能力下降的重要原因。此外,在带毒虫主动取食波时间延长的情况下,被动取食行为减少,休息波次数减少,进一步表明RSMV侵染不利于电光叶蝉的取食,导致其增加主动取食行为,并减少了休息频率。
然而,植物病毒也可以通过间接作用改变介体昆虫的寄主选择倾向性,以利于病毒的传播和扩散(施艳等, 2013)。本研究发现感染RSMV的水稻显著吸引电光叶蝉前来取食,而带毒电光叶蝉倾向于取食健康水稻,表明RSMV对电光叶蝉寄主选择倾向性的影响,有利于病毒在植物间的扩散。该结果与SRBSDV对介体白背飞虱寄主选择倾向性的影响,黄瓜花叶病毒(cucumber mosaic virus, CMV)对介体蚜虫寄主选择倾向性的影响(Maucketal., 2010; Wangetal., 2014)相似,均表现出感病植株对无毒介体昆虫的吸引,以及带毒介体昆虫对健康植株的喜好。表明这种病毒对介体昆虫寄主选择倾向性的影响模式一方面提高了介体昆虫种群的获毒率,另一方面促进了带毒种群对病毒的传播效率,同时也表明持久传播和非持久传播的植物病毒与介体昆虫间存在相似的互作模式。已有的研究表明CMV通过沉默抑制子2b蛋白与拟南芥Arabidopsisthaliana和烟草茉莉酸防御途径下游的JAZ蛋白互作而抑制茉莉酸的生成,从而提高感病植株对介体蚜虫的吸引力 (Wuetal., 2017),而关于RSMV如何调控植物激素或挥发物影响昆虫寄主选择倾向性的机制尚不清楚,后续将进一步深入研究。
