水稻施硅对白背飞虱刺吸和寄主选择行为的影响
2020-03-30贾路瑶刘丹丹侯茂林
贾路瑶, 刘丹丹, 侯茂林
(中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京100193)
水稻Oryzasativa是我国三大主粮之一,其产量占粮食总产量的30%~35%(杨有才和周清明, 2003)。然而,水稻受到100多种害虫的为害(Pathak and Khan, 1994)。白背飞虱Sogatellafurcifera是我国重要的水稻害虫之一,具有迁飞性、毁灭性和暴发性(程家安等, 2008)。白背飞虱通过刺吸韧皮部汁液对水稻植株造成直接伤害(娄永根和程家安, 2011),还可以通过传播植物病毒对水稻造成间接伤害(周国辉等, 2010)。化学杀虫剂虽然能在很大程度上可有效控制白背飞虱的危害,但长期、大量、不合理地使用农药带来了许多问题。水稻抗虫性是防治白背飞虱经济有效的措施之一,然而生产上可供利用的抗性资源很少。采用栽培管理措施来增强植物的抗虫性成为害虫防治的可供选择方法之一。
硅通常不被认为是植物生长所必需的元素,但由于它在植物营养方面的重要作用,被认为是一种“有益物质”或“准必需物质”(Ma, 2004)。在许多植物中发现,施硅可增强植物对刺吸式和咀嚼式口器昆虫的抗性。已有报道,施硅能够增强水稻对二化螟Chilosuppressalis(韩永强等, 2010; Hou and Han, 2010)、三化螟Tryporyzaincertulas(Savantetal., 1999)、稻纵卷叶螟Cnaphalocrocismedinalis(Hanetal., 2016)、褐飞虱Nilaparvatalugens(Yangetal., 2017a)、白背飞虱(Salim and Saxena, 1992)等的抗性。硅对植物抗虫性的调节作用,一方面为组成性防御机制,即植物组织中硅沉积增加其硬度和耐磨度,降低植物可消化性(Massey and Hartley, 2009; Hanetal., 2015),阻碍昆虫取食;另一方面为诱导性防御机制,包括增强多种保护性酶、抗虫次生代谢物合成酶等的活性(Alagaretal., 2010; Hanetal., 2016)、诱导胼胝质沉积(Yangetal., 2018)和增强挥发性化合物来吸引捕食性和寄生性天敌(Maleck and Dietrich, 1999)。虽然前人报道施硅增强水稻对白背飞虱的抗性(Salim and Saxena, 1992),然而施硅调节水稻对白背飞虱抗虫性的机制尚不明确。本研究旨在分析水稻施硅对白背飞虱刺吸行为和寄主选择行为的影响,以期阐明施硅增强水稻对白背飞虱的抗性的部分机制,为硅肥用于白背飞虱种群调控奠定理论基础。
1 材料与方法
1.1 供试虫源和水稻
白背飞虱初始种群采自农业部桂林作物有害生物科学观测实验站,已在人工气候箱内连续饲养5年以上。
试验中的水稻品种为台中1号(Taichung Native 1, TN1)。水稻种子在温水中浸泡24 h、催芽至露白后,以塑料育秧盒作为苗床播种。一周后移栽到温室笼罩中的塑料桶(直径17 cm,高20 cm)中,每桶4穴,每穴4株。塑料桶内装同一批烘干的砂质壤土,每桶3.5 kg干土。
根据土壤有效硅含量和水稻对硅的需求量,使用Na2SiO3·9H2O作为硅肥。参照Han等(2016)方法设置不施硅、中硅和高硅3种施硅水平: 0(对照), 0.16和0.32 g SiO2/kg土壤。各处理按照每千克土壤施用纯氮0.34 g(包括磷酸氢二铵和尿素中的氮;尿素,含N≥46.4%)、五氧化二磷0.41 g(磷酸氢二铵,含N 21.2%和P2O553.8%)、氯化钾0.62 g(含K2O≥60%)。肥料于移栽前3 d施用,施用后加水搅拌均匀。
1.2 施硅后水稻叶鞘硅化程度的测定
随机选取上述3种处理的35日龄盆栽水稻,在每株稻苗主茎中下部叶鞘上系一个封口膜小袋(5 cm×5 cm),每小袋内接入5头白背飞虱初羽化雌成虫。接虫时使小袋撑开,以便飞虱有足够的空间活动和取食。接虫0, 24, 48, 72 和96 h后移除虫子和小袋,用刀片切取白背飞虱取食部位的叶鞘0.3~0.5 cm。每处理重复3次。试验在人工气候箱(温度27±1℃,相对湿度70%±5%,光周期14L∶10D)中进行。参照杨秉耀等(2006)的方法制备电镜观察样品。水稻叶鞘经4%戊二醛固定48 h, 1%四氧化锇固定3 h, 0.1 mol/L磷酸缓冲液(pH 7.4)浸洗,50%, 70%, 80%, 90%和100%乙醇梯度脱水,再采用CO2临界点干燥法进行脱水处理,使用离子溅射仪(IB-5, Eiko, 日本)镀膜后,用扫描电镜(Regulus8100, Hitachi, 日本)观察水稻叶鞘中的硅细胞数量和大小。在每个处理中,随机选择5个180×扫描电镜图片(0.35 mm2)用于观测硅细胞数量;用Image-Pro Plus图像分析软件(Version 6.0, Georgia, 美国)测定单个硅细胞的面积,每处理重复5次。
1.3 水稻施硅后白背飞虱若虫刺吸行为的测定
采用刺吸电位(electrical penetration graph, EPG)技术记录白背飞虱若虫在不同处理水稻上的刺吸行为,参照Yang等(2017b)方法进行试验。在法拉第笼中使用Giga-8 DC-EPG(Wageningen Agricultural University, 荷兰)记录白背飞虱的刺吸行为。取饥饿1 h的白背飞虱4-5龄若虫,用真空抽气泵(QC-1S,北京市劳动保护科学研究所)进气口端的吸头吸住虫体腹部,将EPG金线(直径18 μm,长3 cm)用导电银胶粘在白背飞虱的前胸背板上。金线的另一端通过长约2 cm左右的铜丝连在铜钉上,将铜钉插入EPG探头。EPG的植物电极为直径2 mm,长10 cm的铜棒,竖直插入水中。将粘有金线的飞虱轻放在水稻叶鞘部位,使其能够自由取食,同时接通EPG电源,开启波形记录软件。通过软件PROBE 3.4(Wageningen Agricultural University, 荷兰)记录并储存刺探波形。每头白背飞虱若虫持续记录8 h,每次记录更换新的水稻和白背飞虱若虫。每处理有效重复数为13次。试验在温度27±1℃,相对湿度70%±5%的安静环境下进行。
1.4 水稻施硅后白背飞虱雌成虫的寄主选择行为
采用笼罩法测定施硅对白背飞虱雌成虫的栖息选择和产卵选择的影响。测定栖息选择性时,选取35日龄盆栽水稻,每种处理各1盆,拔除枯叶和多余的稻苗,每桶保留10株。不同处理稻苗等距离排列在一个圆上(直径20 cm),盆与盆之间大约间隔20 cm。用80目防虫笼(长×宽×高=60 cm×60 cm×60 cm)罩住。在圆心处用培养皿(直径9 cm,高2 cm)接入35头长翅型白背飞虱雌成虫。72 h后统计各处理水稻上的虫数。一个笼罩为一次重复,共重复12次。重复之间轮换笼罩内各处理稻苗的相对位置。测定产卵选择的方法基本相同,不同的是每处理采用3株30日龄稻苗移栽到塑料杯(直径8 cm,高14 cm)中,在防虫笼中心接入10头初怀卵雌成虫,1周后计数各稻株上的若虫数,并剪取稻株于解剖镜下解剖记录未孵化卵数。产卵量为若虫数加未孵化卵数。共重复6次。试验在温控室进行(温度为27±3℃,相对湿度70%±5%)。
1.5 数据分析
百分比数据进行平方根反正弦转换,以满足方差分析的要求。使用SPSS 24.0双因素方差分析(Two-way ANOVA)检验硅处理、白背飞虱为害以及两者交互作用对水稻叶鞘硅化程度影响的显著性,采用单因素方差分析(One-way ANOVA)检验硅处理对白背飞虱刺吸行为和寄主选择影响的显著性,用Tukey氏检验法比较不同处理间的差异显著性。
2 结果
2.1 施硅对水稻叶鞘硅化程度的影响
电镜观察发现,水稻叶鞘中硅细胞呈哑铃形,成对排列(图1: A, B)。双因素方差分析结果表明,硅处理、白背飞虱取食以及两者交互作用对水稻叶鞘硅化程度(硅细胞数和面积)均有显著影响(表1)。白背飞虱取食不同时间时,水稻硅细胞数随施硅量增大而顺次显著增多(P<0.001)(图2: A);硅细胞面积则相反,随施硅量的增大而顺次显著减小(P<0.001)(图2: B)。在不同施硅量的水稻中,白背飞虱取食24 h时的水稻硅细胞数显著多于取食0 h时(P=0.001)(图2: C),而取食72 h时的水稻硅细胞数显著少于取食0 h时(P=0.002)(图2: C);白背飞虱取食0 h时的水稻硅细胞面积显著小于其他取食时间(P≤0.002)(图2: D),取食0 h时比取食96 h时水稻硅细胞面积低6.9%。
2.2 水稻施硅对白背飞虱若虫刺吸行为的影响
EPG记录白背飞虱4-5龄若虫在水稻上的刺吸电位如图1(C)所示。硅处理对非刺探(np)波、路径(Nc)波、韧皮部取食(N4-b)波的总持续时间有显著影响,而且对np波、韧皮部刺穿(N4-a)波、N4-b波的单次持续时间有显著影响(表2)。np波和Nc波的总持续时间随施硅量增大而延长,高硅处理组显著长于对照组(np波:P=0.004; Nc波:P=0.015);而N4-b波的总持续时间随施硅量增大而缩短,施硅处理显著短于对照组(P≤0.041)。np波单次持续时间随施硅量增大而延长,高硅处理组显著长于对照组(P=0.050);与之相反,N4-a波和N4-b波的单次持续时间随施硅量的增大而缩短,高硅处理N4-a波单次持续时间比对照短(P=0.043),施硅处理N4-b波单次持续时间显著短于对照(P≤0.028)。
2.3 水稻施硅对白背飞虱雌成虫寄主选择性的影响
施硅显著影响白背飞虱雌成虫对水稻的栖息选择性(F=244.393,df=2,35,P<0.001)(图3: A)和产卵选择性(F=28.924,df=2,17,P<0.001)(图3: B)。白背飞虱雌成虫在不同处理水稻上的栖息率随施硅量的增大而显著下降(P<0.001)(图3: A),中硅和高硅处理分别比对照降低48.0%和67.4%,高硅比中硅处理降低37.4%。就产卵选择性而言,中硅和高硅处理水稻上的产卵量均显著低于对照(P<0.001)(图3: B),比对照分别降低34.8%和46.1%。
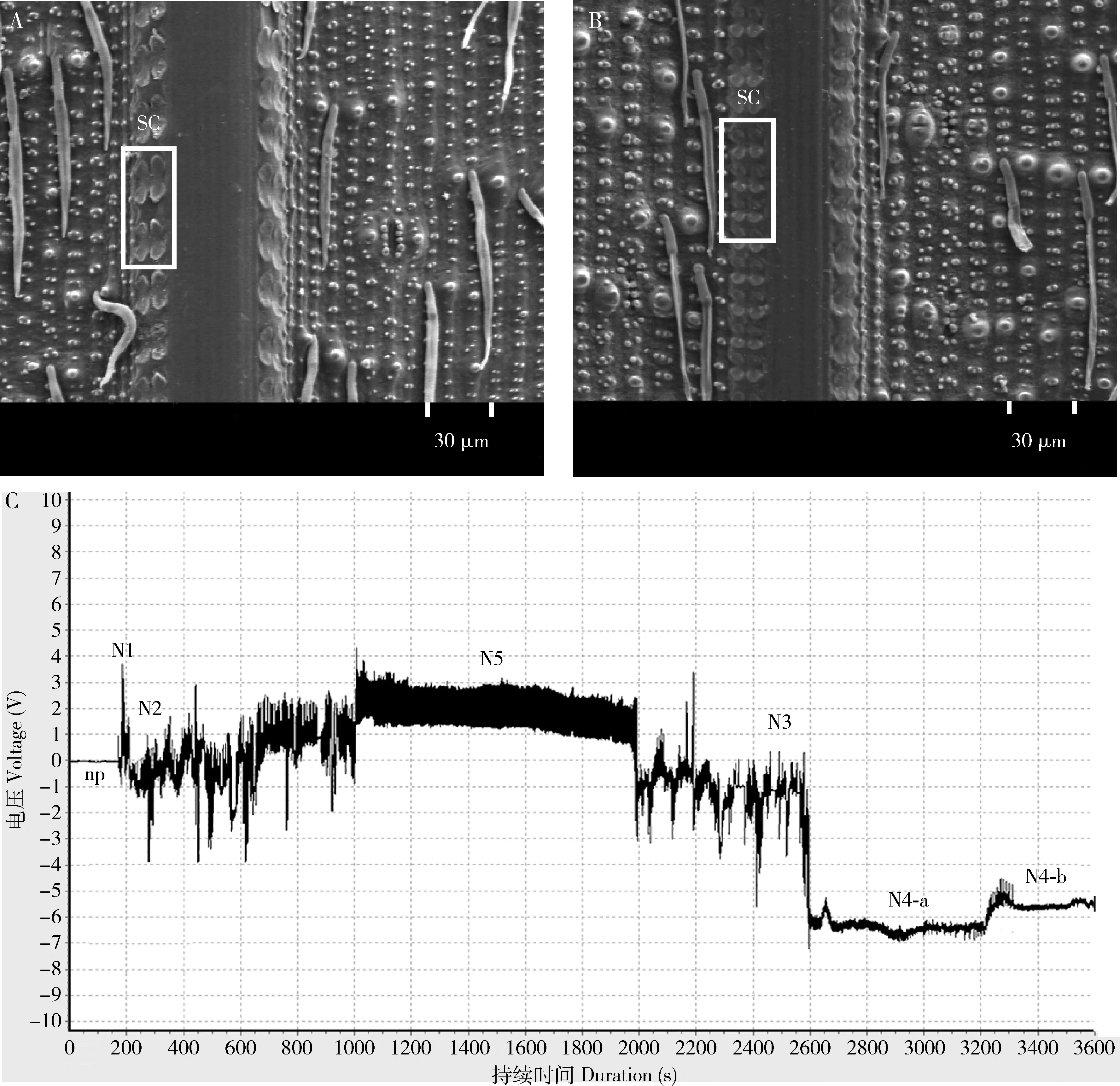
图1 水稻叶鞘表面电镜图和白背飞虱刺吸电位图Fig. 1 Scanning electron microscope (SEM) photos of rice leaf sheath surface and electrical penetration graph (EPG) waveforms identified from Sogatella furcifera on riceA: 对照叶鞘电镜图SEM graph of leaf sheath of the control plant; B: 施硅处理叶鞘电镜图SEM graph of silicon-treated leaf sheath; C: EPG. SC: 硅细胞Silica cell. np: 未刺探Non-penetration; N1: 刺探开始Penetration initiation; N2: 唾液分泌和口针移动Salivation and stylet movement; N3: 口针在韧皮部外活动Extracellular stylet activity near the phloem region; N4-a: 口针刺入韧皮部Intracellular stylet activity in the phloem region without ingestion; N4-b: 取食韧皮部汁液Phloem sap ingestion; N5: 口针在木质部活动Activity in the xylem region. 其中,N1, N2和N3合并为路径(Nc)波N1, N2 and N3 are collectively named as pathway event (Nc).

表1 硅处理和白背飞虱为害影响水稻叶鞘硅化程度的双因子方差分析参数Table 1 Parameters of two-way analysis of variance for significance of the effects of silicon amendment and Sogatella furcifera infestation on silicification of rice leaf sheaths
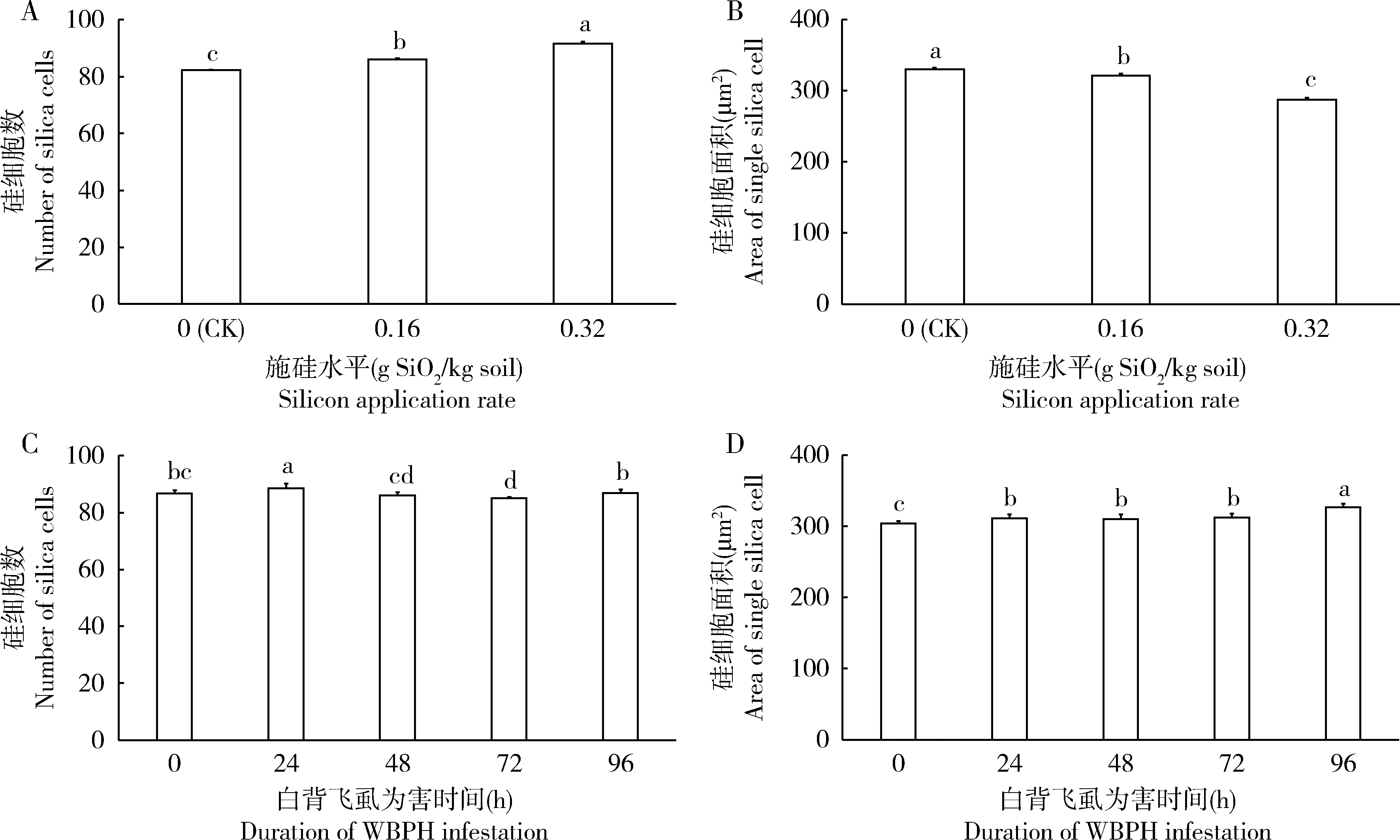
图2 硅处理和白背飞虱为害影响水稻叶鞘硅化程度的主效应Fig. 2 Main effects of silicon application and Sogatella furcifera infestation on silicification of rice leaf sheaths柱上不同字母表示处理间差异显著(Turkey氏检验, P<0.05)。Different letters above bars indicate significant difference (Turkey’s test, P<0.05).

表2 EPG记录的施硅和未施硅水稻上白背飞虱若虫的取食行为Table 2 Feeding behaviors of Sogatella furcifera nymphs on rice plants amended with Si or not recorded by EPG

图3 白背飞虱雌成虫对施硅水稻的栖息(A)和产卵(B)选择性Fig. 3 Settling (A) and oviposition (B) preference of female adults of Sogatella furcifera on rice plants amended with siliconCK: 不施硅No silicon was amended. 数据表示为平均值±标准误,柱上不同字母表示差异显著(Tukey氏检验, P<0.05)。Data are expressed as mean±SE. Bars with different letters indicate significant difference (Tukey’s test, P<0.05).
3 讨论
植物抗虫机制表现为抗生性(antibiosis)、耐害性(tolerance)和排驱性(antixenosis)3个方面(Kochetal., 2016)。排驱性是指植物使昆虫不趋向其上取食、产卵或栖息的一些特性。植物的表面形态、生理生化特性和分泌的挥发性次生物质可以阻止昆虫趋向植物产卵或取食,从而避免或减轻害虫为害(卢伟等, 2007; 韩永强等, 2017)。水稻对白背飞虱的抗性与水稻叶鞘表皮的刚毛、刺毛、硅细胞群、硅细胞链、木栓层和蜡被层密度等密切相关(叶海芳, 1989)。本研究结果表明,白背飞虱雌成虫对中硅和高硅处理水稻的栖息选择率(图3: A)和产卵选择性(图3: B)均显著低于对照,表明施硅增强了水稻对白背飞虱的排驱性,这可能与施硅增大水稻叶鞘的硅化程度有关。水稻是典型的硅酸植物,具有主动吸收和富集硅的能力(Maetal., 2006)。吸收的硅富集在水稻茎秆、叶鞘和叶片的表皮,增加植物组织的硬度和耐磨度。本研究结果(图2)和此前的报道(Yangetal., 2017b)均发现施硅增加单位面积水稻叶鞘上的硅细胞数量,这有助于增强表皮上的刚毛、刺毛、木栓层和蜡被层对白背飞虱寄主选择的不利影响,从而导致白背飞虱对施硅水稻的栖息和产卵选择性下降。另外,Kvedaras等(2010)报道施硅影响植物挥发性次生物质的释放,但施硅是否影响水稻挥发性次生物质的释放及其在施硅水稻对白背飞虱的排驱性中的作用,需要开展进一步的研究。
刺吸式害虫通过口针从植物韧皮部汁液获取营养。在刺吸植物韧皮部汁液之前,口针需要经过一系列的刺探行为(Leietal., 2016)。在这个过程中,植物的组织化学特性会对刺吸式害虫的刺吸行为产生影响。本研究中,我们发现施硅影响白背飞虱若虫对水稻韧皮部汁液的摄取,施硅处理缩短了白背飞虱若虫的韧皮部取食(N4-b)的时间(表2);这与施硅对褐飞虱(Yangetal., 2017b)和麦二叉蚜Schizaphisgraminum(Costaetal., 2011)刺吸行为的影响一致。有意思的是,这一结果也与褐飞虱在抗虫品种上的行为相似,即褐飞虱在携带抗性基因Bph14和Bph15的水稻植株上摄取韧皮部汁液的时间减少(Haoetal., 2008)。在施硅处理水稻上,白背飞虱的非刺探(np)波的总持续时间和单次持续时间以及路径(Nc)波的总持续时间均显著高于对照水稻(表2)。再者,虽然韧皮部刺穿(N4-a)的总持续时间在施硅处理与对照之间没有差异,但是对照中单次持续时间反而显著长于施硅处理(表2),这是由于白背飞虱在施硅水稻上进行了更为频繁的刺探和刺穿行为。这些结果表明白背飞虱在施硅水稻上的刺吸行为受到了阻碍。扫描电镜发现,施硅水稻的硅化程度增大,而且硅处理的水稻叶鞘维管束四周有排列紧密的硅细胞链,这可能使得白背飞虱在施硅水稻上花费更多的时间来选择刺探部位(即np波持续时间增大),而且白背飞虱在施硅水稻上选择的刺探部位可能并不适合进一步的韧皮部刺穿,从而表现出更频繁的刺探和刺穿。硅处理水稻上韧皮部取食(N4-b)时间的下降,一方面可能是施硅使得水稻韧皮部汁液中可溶性硅酸含量上升(这需要进一步测定),因为水稻叶鞘中提取出的可溶性硅酸可以减少褐飞虱取食(Sowers and Thurston, 1979);另一方面,施硅处理可能参与白背飞虱刺吸诱导的胼胝质在韧皮部筛管上的沉积(Yangetal., 2018),从而阻止白背飞虱取食韧皮部汁液。胼胝质是一种植物多糖,虫害可诱导其在韧皮部筛管上沉积,从而阻止刺吸式害虫的韧皮部汁液取食(Haoetal., 2008)。施硅水稻上白背飞虱刺吸行为受阻将最终导致白背飞虱取食量下降,从而增强施硅水稻对白背飞虱的抗生性。
本研究结果表明,施硅增强水稻对白背飞虱的排驱性、阻碍白背飞虱的刺吸行为,这将有助于降低施硅水稻上白背飞虱的种群和为害,从而增强水稻对白背飞虱的抗性。
