封育年限对毛竹林群落结构和林下植物多样性的影响
2020-03-27叶柳欣库伟鹏徐旻昱孟方荣傅伟军吴家森
叶柳欣,库伟鹏,刘 军,徐旻昱,孟方荣,傅伟军,刘 娟,金 锦,吴家森,*
1 省部共建亚热带森林培育国家重点实验室, 临安 311300 2 浙江农林大学环境与资源学院, 临安 311300 3 杭州市余杭区林业工作站, 余杭 311100 4 杭州格真农产品专业合作社, 余杭 311118
森林群落的林分密度、乔木树种的胸径分布和林下植物多样性反映了森林群落的形成、稳定性及演替规律,影响着森林生态功能的发挥和立地生产力的可持续性[1- 3]。不同林龄人工林林下植物种类多样性预示着生态系统的稳定性,标志着人工林演替的方向和进程[4]。
毛竹是我国重要的森林资源,广泛分布于我国南方的浙江、福建、江西、湖南等省份,面积达443×104hm2,具有生长快、产量高、用途广、可持续性强等特点[5- 6],2016年,浙江省毛竹林面积80.71×104hm2,总株数297888万株[7]。毛竹林群落结构及其稳定性受到人为活动的强烈支配和制约,表现为不同干扰程度的毛竹林乔木层结构和林下植物多样性有显著差异。研究表明,材用毛竹林胸径大于同等管理措施下的笋用林[8];长期经营后毛竹林群落结构简化,林下灌木多样性降低[9- 10];随着人为经营强度的增大,毛竹林下植物的多样性、丰富度指数均随之减少[11- 12];也有研究认为人为经营后毛竹林下灌木、草本的物种多样性上升,草本层多样性指数、均匀度指数及物种数均以重度干扰的毛竹林为最高[13]。
随着我国社会、经济的发展,毛竹价格逐年下降[14],劳动力成本持续上升[15],导致毛竹砍伐人工成本大于竹材销售收入,造成了浙江省较大面积的集约经营毛竹林疏于管理,处于自然封育状态。此外,由于经济发展过程中的人口城市化迁移,大量青壮年人口涌入经济发达地区,致使欠发达山区的青壮年劳动力水平下降,加剧了农村人口老龄化问题[16],在一定程度上也会造成毛竹林的荒芜。自然封育后毛竹林的群落结构和林下植物的物种组成、多样性如何演变,至今还未见报道。鉴于此,本研究以浙江省典型毛竹纯林为研究对象,采用时间代替空间的方法,通过建立标准地全面调查和研究了不同封育年限毛竹林的胸径分布和林下植被多样性,为毛竹林的群落演替和生态功能的评价提供基础和理论依据。
1 材料与方法
1.1 研究区概况
研究区位于有“中国竹子之乡”之称的浙江省杭州市余杭区境内,地理坐标为E119°42′2.71″—119°50′35.22″、N30°25′28.1″—30°33′34″。属于北亚热带南缘季风气候区,冬夏长春秋短,温暖湿润,四季分明,光照充足,雨量充沛,年平均气温15.3—16.2℃,年平均降雨量1391.8mm,年平均相对湿度76%,无霜期199—328d。
1.2 试验设计与研究方法
2017年7月,通过查阅杭州市余杭区森林资源经营档案和全面踏查的基础上,选择余杭西部的百丈镇和鸬鸟镇封育年限分别为10年、20年和30年的毛竹纯林作为研究对象,以常规经营的竹林为对照(0年),设置不同封育年限样地各9个,共计36个样地,样地面积为30m×30m。封育毛竹林常年没有人为活动干扰,没有挖笋、清理老竹等活动,完全处于自然封育状态。常规经营毛竹林每年5 月上、中旬施肥1 次,施用600 kg/hm2的复合肥(m(N)∶m(P2O5)∶m(K2O)为15∶15∶15),肥料地表撒施,结合松土垦复15 cm左右。常年进行挖笋活动(春笋、鞭笋、冬笋),毛竹砍伐集中在小年。样地基本情况如表1所示。
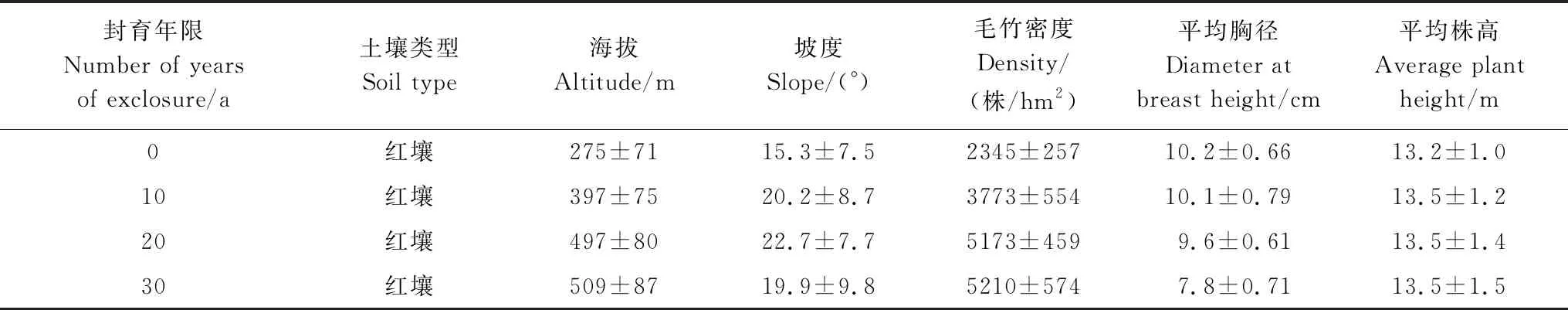
表1 不同封育年限毛竹林样地基本情况
全面测量样地中不同林龄(1度竹、2度以上活立竹)毛竹和枯立竹的胸径,调查林下灌木、草本植物的种类、株数、高度、冠幅及盖度[17-18]。
1.3 数据处理
根据样地调查结果,计算各封育年限不同林龄毛竹林密度、胸径;同时计算灌、草重要值、Shannon-Wiener指数、Simpson指数、Pielou指数[19]。
灌草重要值=(相对多度+相对盖度+相对频度)/3
Jsw=H′/lnS
H′为Shannon-Wiener指数,Ds为Simpson指数,Jsw为Pielou指数,式中Pi=ni/N,Pi为第i种植物在样地中出现的概率,N为样地中植物总个体数,S为总种数,ni为第i种植物个体数。
本文数据整理使用Excel 97-03,显著性差异分析使用SPSS 18.0,制图使用Origin 8.5。
2 结果与分析
2.1 毛竹林群落结构
毛竹林中不同林龄毛竹和枯立竹的密度,一定程度上反映了毛竹林的结构。由图1可知,不同封育年限下毛竹林1度竹的平均密度为727—824株/hm2,它们之间没有显著差异(P>0.05);封育条件(10年、20年、30年)下毛竹林中2度以上活立竹的平均密度显著高于常规经营(0年),封育为20、30年的竹林密度又显著高于10年(P<0.05)。枯立竹的密度随着封育年限的延长而显著增加(P<0.05)。
胸径是竹林群落结构的重要指标,是毛竹的生长状况及土壤养分供给的表现。由表1和图1可知,随着自然封育年限的延长,毛竹胸径呈现下降的趋势。封育30年的1度竹的平均胸径显著低于其他年限(P<0.05)。2度以上活立竹则表现为0年、10年显著大于20年(P<0.05),同时也显著高于30年(P<0.05)。枯立竹则表现为10年>20年>30年,不同封育年限的差异达显著水平(P<0.05)。
不同林龄毛竹和枯立竹在各封育年限下胸径的分布如图2所示,从图2中可知,封育年限为0—20年的1度竹胸径分布服从负偏态分布,峰值向右偏移,封育30年的1度竹胸径分布则大体服从正态分布,峰值居中。2度以上活立竹的胸径分布则表现为封育年限为0年和10年呈现负偏态分布,封育20年表现为正态分布,封育30年服从正偏态分布(图2)。枯立竹的胸径分布则表现为封育10年和20年的竹林呈现正态分布,封育30年表现为正偏态分布。随着封育年限的增加,胸径分布的峰值会从右往左移动,即小胸径枯立竹所占比例增加(图2)。
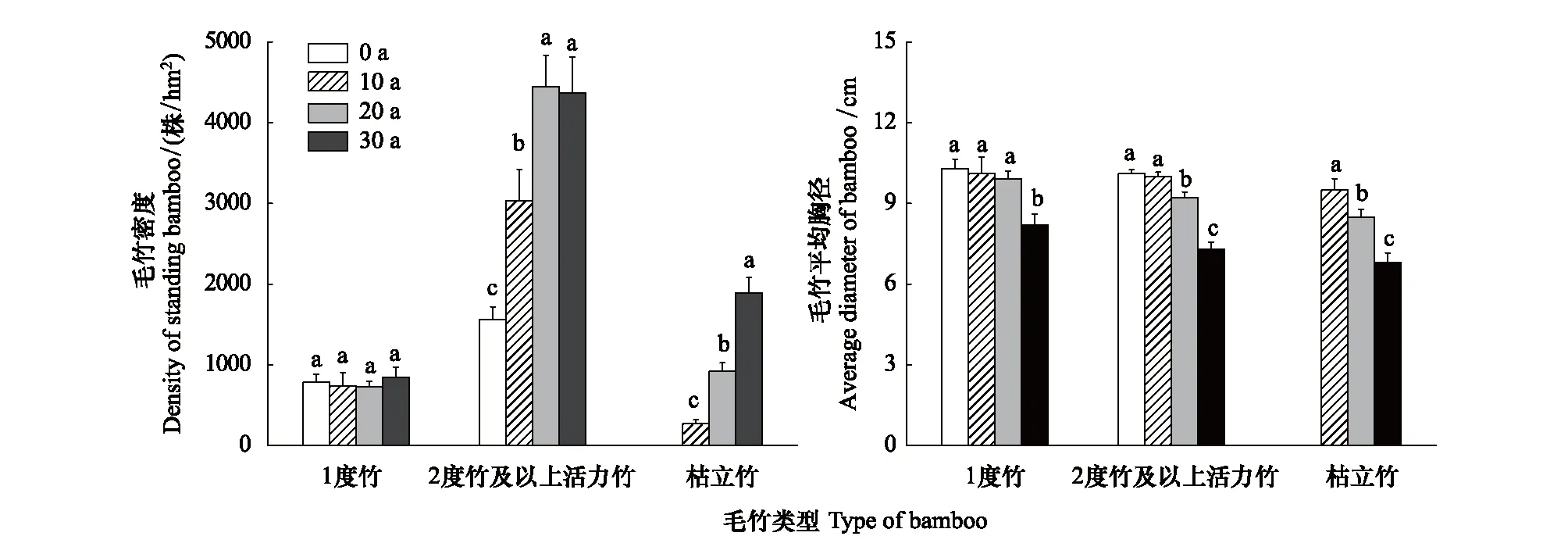
图1 不同封育年限下毛竹密度和平均胸径Fig.1 Density and diameter at breast height of bamboo in different natural exclosure years
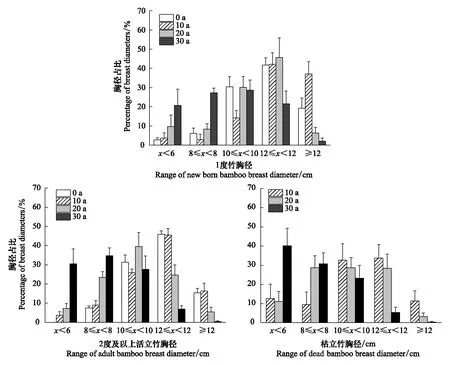
图2 不同封育年龄下毛竹胸径分布情况Fig.2 Distribution of diameter at breast height of bamboo in different natural exclosure years
2.2 林下灌木物种组成及重要值
植物的重要值可以用综合数值来表现该植物物种在群落中的相对重要性,是评估物种多样性的重要指标。不同封育年限毛竹林下主要灌木植物(重要值≥1)物种组成及重要值如表2所示,所有毛竹林中均出现的灌木植物有山莓、络石、紫金牛、山橿、青灰叶下珠和小果菝葜。封育年限为0年时,重要值排名前5的灌木植物依次为山莓、茶、寒莓、络石、紫金牛,其中山莓(12.777%)和茶(12.029%)是占优势地位的灌木物种。封育年限为10年的竹林下则依次为络石、山橿、茶、毛花连蕊茶、紫金牛,其中络石(22.829%)和山橿(19.775%)的重要值明显高于其他灌木植物。20年依次为紫金牛、牛鼻栓、山橿、马银花、异叶爬山虎,其中紫金牛(10.880%)、牛鼻栓(10.457%)和山橿(9.851%)是毛竹林下主要的优势灌木。30年依次为山橿、牛鼻栓、茶、蓬莱葛、寒莓,其中山橿(11.702%)的优势度明显。
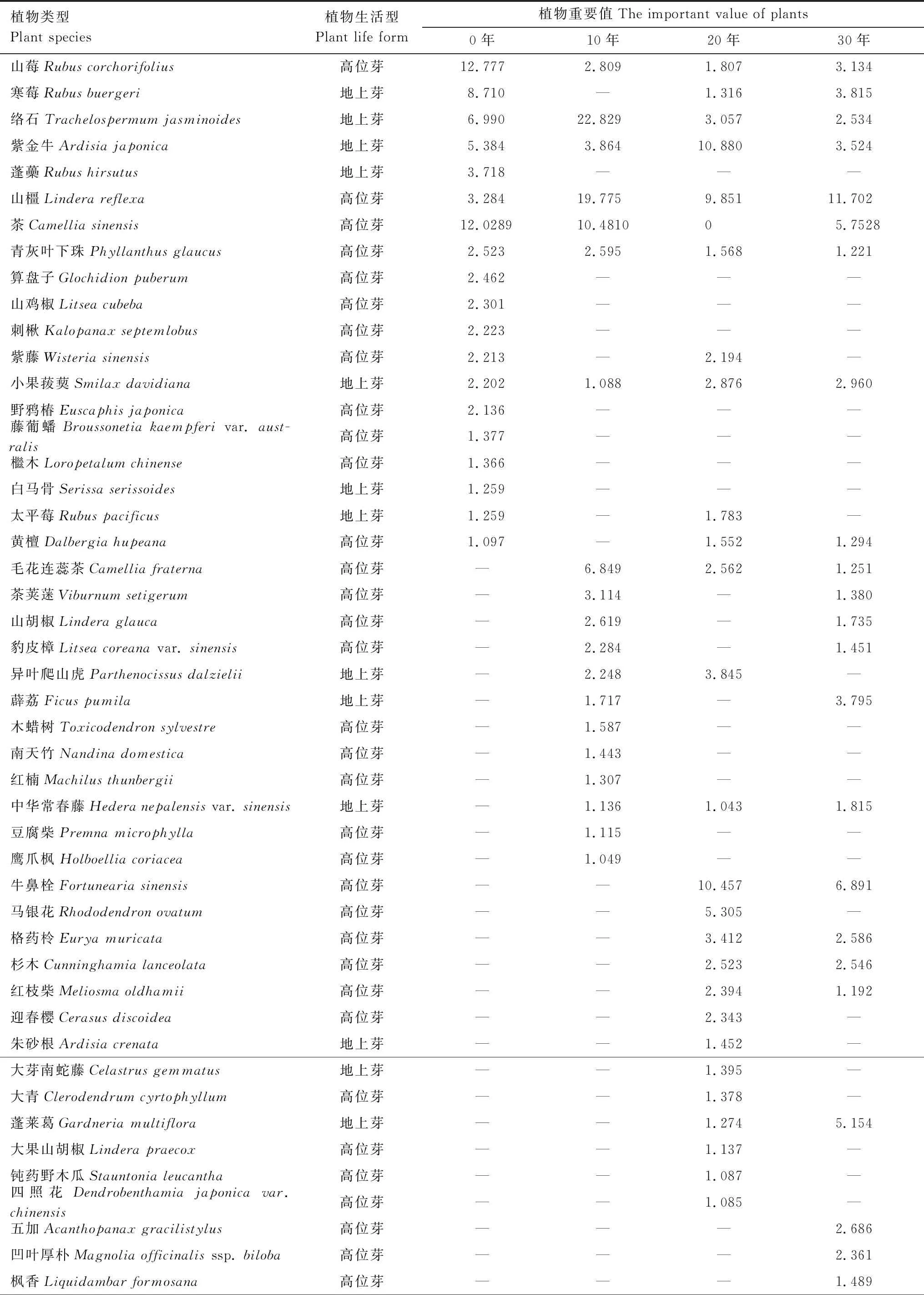
表2 不同封育年限毛竹林下主要灌木植物的重要值
2.3 林下草本植物种类及重要值
不同封育年限毛竹林下主要草本植物(重要值≥1)物种及重要值如表3所示,所有封育年限下均有分布的草本植物有褐果薹草、求米草、华东蹄盖蕨、玉竹、阔叶箬竹和金星蕨。封育年限为0年时,重要值前5的草本依次为金毛耳草、疏羽凸轴蕨、淡竹叶、褐果薹草、求米草;10年时依次为麦冬、褐果薹草、华东蹄盖蕨、条穗薹草、紫萁;20年时则为求米草、阔叶箬竹、华东蹄盖蕨、条穗薹草、麦冬;30年依次为阔叶箬竹、绵穗苏、玉竹、山麦冬、华东蹄盖蕨。常规经营条件下(0年),金毛耳草(11.999%)和疏羽凸轴蕨(9.252%)的重要值明显大于其他草本。封育为10年时,麦冬(15.029%)、褐果薹草(10.931%)、华东蹄盖蕨(10.785%)和条穗薹草(10.649%)在竹林下都占据一定重要值。随着封育的进行,20年时求米草(20.159%)、阔叶箬竹(16.065%)和华东蹄盖蕨(11.234%)的重要值增大到一定水平。封育为30年时,阔叶箬竹(37.224%)成为林下绝对优势草本。
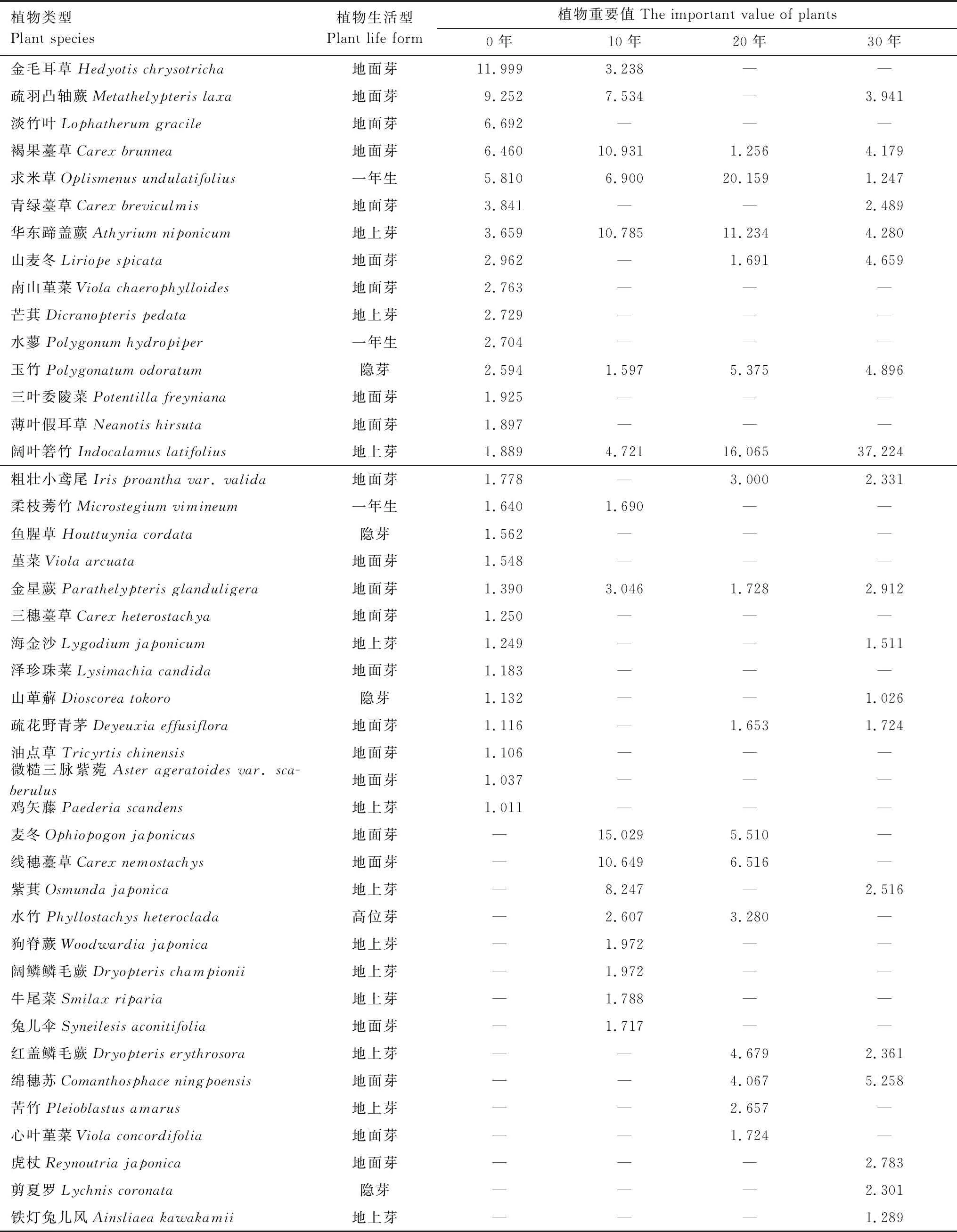
表3 不同封育年限下主要草本植物的重要值
2.4 林下灌木和草本多样性指数
不同封育年限毛竹林下灌木的优势度指数Simpson、多样性指数Shannon-Wiener和均匀度指数Pielou如表4所示。从表中可知,封育10年时的毛竹林下灌木的Simpson指数显著低于其他林分(P<0.05);灌木Shannon-Wiener指数则表现为20年和30年的林分显著高于10年和0年(P<0.05), Pielou指数则表现为0年显著低于10年、20年和30年的林分(P<0.05),10年、20年和30年间数值存在波动,峰值出现在20年。
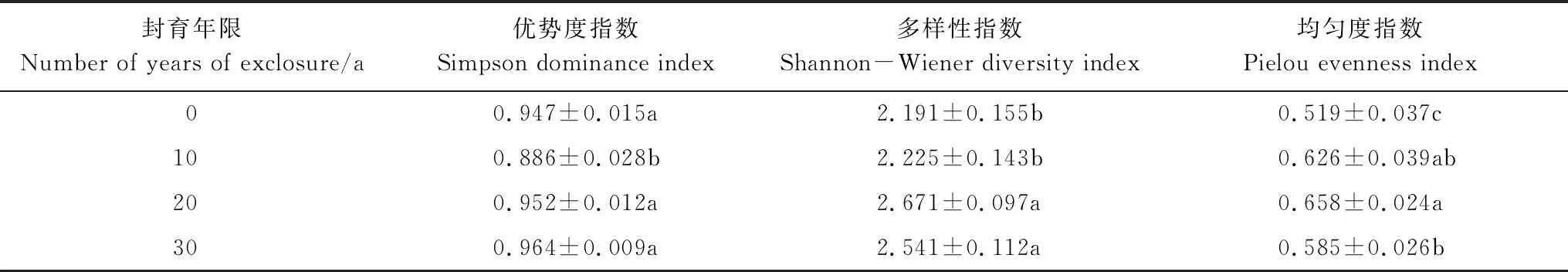
表4 灌木植物多样性指数
从表5可知,随着封育年限增长,草本植物的Simpson指数呈现下降趋势,0年的优势度指数显著高于10年和20年(P<0.05),同时显著高于30年(P<0.05)。多样性指数Shannon-Wiener表现为常规经营(0年)的林分显著高于封育20年以后的林分(P<0.05),而不同封育年限间的差异不显著。自然封育10年草本的均匀度指数Pielou显著高于其他林分(P<0.05),且封育为0年和20年林分中的草本均匀度指数Pielou又显著高于自然封育为30年的林分(P<0.05)。
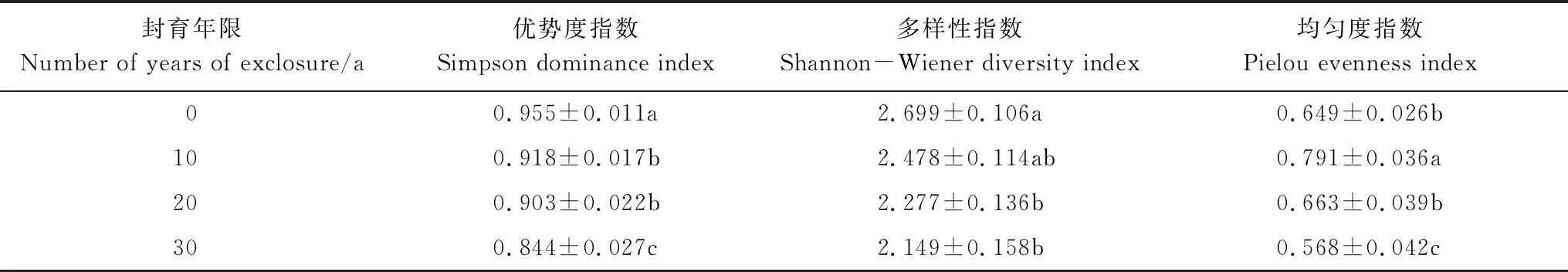
表5 草本植物多样性指数
3 讨论与结论
3.1 自然封育对毛竹林分结构的影响
毛竹林总密度随着封育年限的延长而增加。在自然封育条件下,单位面积毛竹林中1度竹株数没有发生明显变化,2度以上活立竹株数逐渐增加,封育20年后基本保持稳定。
1度竹株数在不同封育年限间没有显著差异。主要原因是常规经营毛竹林样地的1度竹受人为选择性留养,一般种竹留养量约800株/hm2。自然封育10年后,毛竹林密度达3000株/hm2,由于水、肥、光等因子的限制,新竹成竹的难度加大,1度竹的密度也基本保持在800株/hm2,随着自然封育时间的进一步延长,老竹逐渐死亡,枯立竹数量大大增加,样地内新生毛竹的数量与死亡毛竹数量基本趋于平衡。
2度以上活立竹逐渐增加的主要原因有:常规经营毛竹林经营中一般遵循“砍四留三”的原则,即砍去4度以上的毛竹,而保留3度及以下的毛竹,除1度竹外,样地中的毛竹为2—3度,密度大约在1600株/hm2。自然封育初期,毛竹生长不再受人为干扰的影响,毛竹林中包括1—6度活立竹,因此其密度明显提高。随着自然封育年限的进一步延长,毛竹林内生存资源开始短缺,造成了2度以上的活立竹增长速率减缓并最终趋于稳定,每年枯死的毛竹也在逐渐增加。
毛竹林平均胸径随封育时间的延长而逐步下降。较小胸径毛竹的株数占毛竹总株的比例逐渐增大,毛竹胸径趋于减小。这与王平等人的研究结果相似,即当立竹密度增加时,毛竹平均胸径趋于减小,毛竹的平均胸径服从自疏法则[20]。在空间和资源的限制下,竞争是导致毛竹平均胸径下降和总密度趋于稳定的主要因素。自然封育20年后毛竹林总密度没有显著差异,但平均胸径则表现为自然封育30年的林分显著低于20年。
3.2 自然封育对毛竹林下植物种类与生活型的影响
毛竹林下植物群落的科属组成复杂,植物种类丰富。其中主要灌草植物(表2—3)隶属50科81属,总共94种,其中被子植物42科72属84种,裸子植物1科1属1种,蕨类植物7科8属9种。封育年限为0,10,20,30年毛竹林下灌木种类分别为68,35,58,77种,而草本植物种类则分别为64,23,31,44种。灌草物种丰富度表现为常规经营>自然封育(年限小于等于20年),这与方茹意等人的研究结果相似,即适度的人为干扰有利于维持林中灌草的物种丰富度[21]。封育条件下灌、草物种丰富度表现为30年>20年>10年,毛竹林下灌木、草本植物种类随封育时间的延长逐渐增加。可见长期的自然封育也对提高竹林灌、草物种丰富度有一定的帮助。
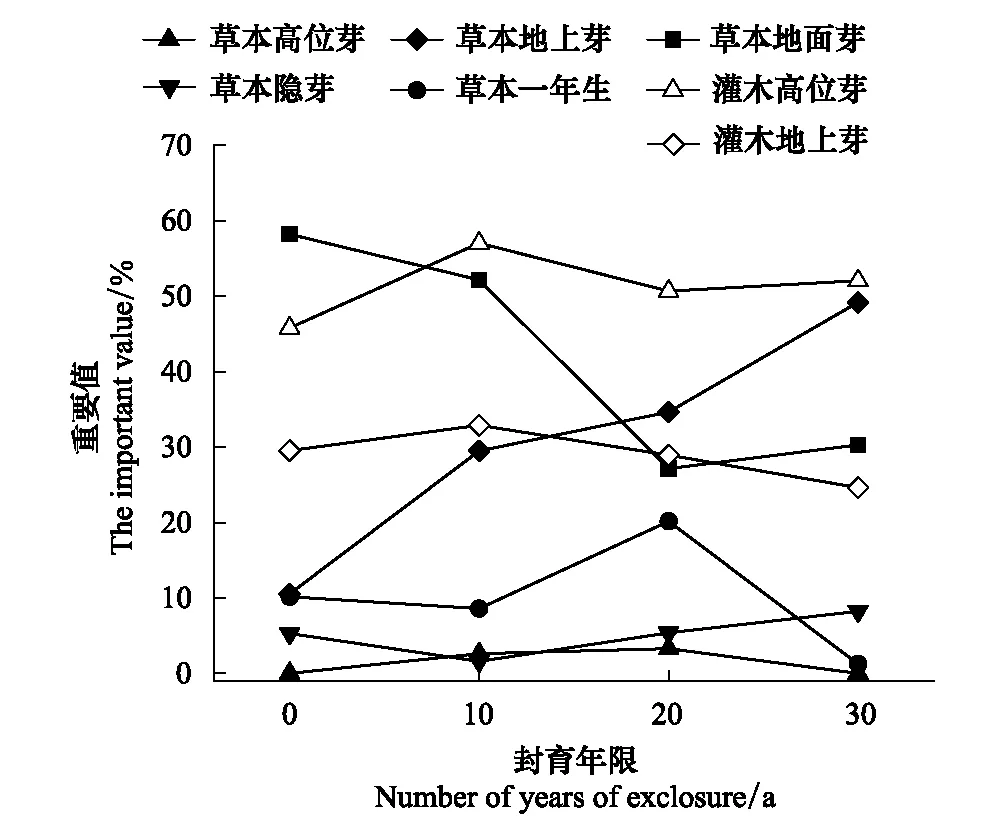
图3 毛竹林下不同生活型主要灌草的重要值变化情况 Fig.3 Changes in important values of main shrubs and herbs under different life forms under bamboo forests
在封育过程中,山橿是一种优势灌木,在各封育年限(10年、20年、30年)下都有较高重要值,而在常规经营(0年)竹林下,山橿的重要值却很低。研究表明山橿属于多年生灌木,生长缓慢,一般生长在山地树林下,喜欢土层深厚,土壤肥沃和半阴凉的环境[22]。毛竹林下草本层中阔叶箬竹随着封育年限的延长,重要值连续升高,封育30年后,其值高达37.224%,是该阶段的绝对优势草本。阔叶箬竹为灌木状草本,径0.5—1.5cm,节间长5—22cm,高1—1.5m,最大竿高可达2m,对土壤要求不严,较耐寒,喜湿耐旱,可在山谷、山麓和山腰地带等迅速生长扩延[23-24],对土壤、温度、水分的要求不高,且作为灌木状草本,其株高、盖度在吸收光线方面占有一定优势,竞争能力极强。因此阔叶箬竹逐渐淘汰其他草本成为长期封育毛竹林下的绝对优势草本。
如表2、3所示,林下主要灌、草植物按照出芽方式分为5种生活型(高位芽、地上芽、地面芽、隐芽和一年生植物),不同封育年限下各生活型重要值之和的变化一定程度上能表现林下灌、草植被在自然封育条件下的演替方向(图3)。
根据图3可知,地面芽草本植物在毛竹林中随封育年限的延长有较为明显的衰退,而地上芽草本植物的重要值则在该过程中显著增长,此外一年生草本植物在封育为30年时几乎绝迹。灌木植物的重要值变化规律相对不明显,表现为高位芽灌木重要值小幅上升,地上芽灌木重要值小幅下降。
植株的出芽高度一定程度上体现了其对光照的竞争能力,自然封育条件下,在林分日渐闭郁的竹林中,出芽较高的植物相对容易接受光照,这点在生态位处于最下层的草本植物中有更为明显的体现。因此,长期的自然封育过程中,由于竹林密度增大,光照逐渐稀缺,拥有易受光和耐阴特性的林下植被更容易在竹林下繁荣。
3.3 自然封育对毛竹林下植物多样性指数的影响
优势度指数Simpson、多样性指数Shannon-Wiener和均匀度指数Pielou等多样性指数可以表征林分的植物多样性情况,体现不同封育阶段林分下灌木层和草本层的植物资源丰富程度。封育为10年的毛竹林分下灌木层的优势度指数Simpson显著低于其他林分。主要是该阶段的林分下存在绝对优势的灌木植物,从表1可知,络石和山橿在封育为10年的林下占据极高的重要值,其值分别为22.829%,19.775%。灌木多样性指数Shannon-Wiener在封育过程中大体表现为先升高后降低,其中以封育20年的林分为最大。随着封育时间的进一步延长,灌木多样性呈现下降趋势,主要原因是由于毛竹密度增加到一定阈值,林分闭郁,林下光线匮乏,植物间的竞争更为激烈,抑制了灌木植物的生长[25-26]。灌木均匀度指数Pielou随封育年限的延长先增大而后降低,0年和封育30年的林下灌木均匀度指数Pielou较低。这可能是由于0年和30年的林下灌木物种丰富度S较高,致使均匀度指数的计算公式Jsw=H′/InS中的分母InS比较大,导致Pielou指数值的下降。这与马克平等人的研究结果相似,即物种的均匀度指数Pielou与多样性指数Shannon-Wiener呈正相关,与物种丰富度S呈负相关[27]。综上,在一定时长范围内,自然封育可以提高毛竹林下灌木植物的多样性,但长期的自然封育,灌木群落多样性会呈现为一定程度的衰退。
草本植物的优势度指数Simpson随封育年限的增加而下降,草本植物阔叶箬竹的优势地位在封育过程中逐渐突出,封育年限为30年时在林下占有绝对优势。多样性指数Shannon-Wiener也随着封育时间的延长而逐渐下降,表明草本植物的多样性水平与封育年限呈负相关。均匀度指数Pielou与封育年限大体呈负相关的关系。随着封育的进行,草本植物群落逐渐衰退,个别草本的优势地位逐渐突出,草本植物的物种多样性水平明显下降。这主要是由于在日渐闭郁的林分中,竹林密度日益增大,生态位处于最下层的草本植物,对光照等资源的反应更为敏感,面临的竞争更为残酷,草本层植物群落衰退,只有少数适应能力强的种类可以在林下大量生长,成为绝对优势的品种[28]。
