艰难梭菌二元毒素A 的原核表达及免疫原性分析
2020-03-22黄颖凤林雪霏黄欢欢吴爱武
黄颖凤,林雪霏,黄欢欢,吴爱武
(广州医科大学金域检验学院,广东 广州 510182)
艰难梭菌是产芽孢的革兰阳性杆菌,属于梭状芽孢杆菌属中的专性厌氧菌[1]。艰难梭菌主要的致病因子为毒素A和毒素B,一部分高产毒的艰难梭菌还可以产生二元毒素(Clostridium difficilebinary toxin,Cdt),该毒素由CdtA和CdtB 2个独立的蛋白亚基组成,其中CdtA具有酶活性,CdtB具有结合能力[2-3],Cdt具有肌动蛋白特异的二磷酸腺苷核糖基转移酶活性,可导致细胞骨架解聚[4],产Cdt的菌株可使毒素A表达量提高到约为普通菌株的16倍,毒素B表达量提高到约为普通菌株的23倍[5],从而产生更强的致病性,导致更高的死亡率。艰难梭菌感染(Clostridium difficileinfection,CDI)主要表现为不同程度的腹泻和伪膜性肠炎[6]。2003年以来,北美、欧洲等地区CDI发病率和病死率明显增加,这与曾暴发流行的RT 027型艰难梭菌以及一小部分RT 078型高产毒CDI有关[5]。除此之外,暴发流行的艰难梭菌菌株还有RT 017、RT 018、RT 106、RT 176和RT 244型。2008年,我国香港首次报道发现高产毒的RT 027型艰难梭菌菌株[7],但目前尚未见全国性CDI流行病学调查分析结果的报道,因欧洲和北美洲高产毒艰难梭菌菌株暴发感染的前车之鉴[8],分泌Cdt的高产毒艰难梭菌菌株之潜在危害不可小觑。目前,抗菌药物治疗CDI复发率高达20%[9],为了防止艰难梭菌引起的大规模CDI暴发,给易感患者接种艰难梭菌疫苗是最理想的方法,该疫苗接种最适用于计划住院治疗、需要长期护理、入住养老院和需要长期或高频率服用抗菌药物的人群,但经过20多年的研究,目前仍无被批准上市的艰难梭菌疫苗[10-11]。本研究以可引起CDI暴发流行的、能产生Cdt的艰难梭菌为研究重点,对CdtA的原核表达及免疫原性进行初步探究。
1 材料和方法
1.1 菌种来源及试剂
pET-28b质粒保存菌种及包被96孔板的CdtA(C129-1392)蛋白由广州医科大学金域检验学院实验室构建并保存;艰难梭菌RT 027型菌株DNA由加拿大圭尔夫大学Scott博士惠赠;大肠埃希菌BL21感受态细胞、小剂量质粒提取试剂盒等购自北京天根生化科技有限公司;限制性内切酶、Premix Taq聚合酶等购自大连TaKaRa公司;羊抗鼠IgG二抗-HRP购自中国台湾arigo公司;二喹啉甲酸(bicinchoninic acid,BCA)蛋白定量试剂盒购自美国Thermo Fisher Scientific公司;聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜、电化学发光(electrochemical luminescence,ECL)曝光试剂盒购自美国Millipore公司;弗氏完全、不完全佐剂购自美国Sigma公司。实验中所用聚合酶链反应(polymerase chain reaction,PCR)引物合成及基因测序由上海英潍捷基有限公司完成。
1.2 实验方法
1.2.1 引物合成 根据GenBank上的cdtA基因全长序列,利用Primer Premier软件设计引物,并在上下游引物5'端分别添加限制性内切酶及其保护碱基。上游引物为5'-CATGCCATGG GC AAAAAATTTAGGAAACATAAAAGGATT-3'(斜体字母为保护碱基,下划线部分为NcoⅠ限制性内切酶,加框部分为补齐碱基),下游引物为5'-GCGCAAGCTTAGGTATCAATGTTTGCA TCAACAATTAA-3'(斜体字母为保护碱基,下划线部分为HindⅢ限制性内切酶)。
1.2.2 PCR扩增cdtA全长序列 以RT 027型艰难梭菌DNA为PCR模板,按大连TaKaRa公司的高保真酶PremixSTAR HS的使用方法设定PCR循环条件,将PCR产物进行1%琼脂糖凝胶电泳分析,初步鉴定扩增结果。
1.2.3 pET-28b-cdtA原核表达载体的构建 纯化后的PCR产物与pET-28b载体分别经NcoⅠ和HindⅢ双酶切后连接,导入大肠埃希菌BL21感受态细胞,涂布于kana/LB/X-Gal平板,随机挑取23个白色菌落,分别增菌后用pET-28b载体的通用引物T7和T7ter行菌液PCR,选取阳性菌株测序。
1.2.4 His-CdtA重组蛋白的表达 取构建成功的pET-28b-cdtA表达菌株大量增菌,诱导蛋白表达,探索可溶性重组蛋白大量表达的最佳条件。当菌液达到对数生长中期[600 nm处吸光度(A)值=0.6]时,摇床温度分别设定为25 ℃、30 ℃、37 ℃;异丙基-β-D硫代半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)浓度分别为0、0.5、1.0和1.5 mmol/L,时间分别为8、10和12 h;同时设立pET-28b空载体为阴性对照,分别收集不同条件下诱导的菌液,1 789×g离心15 min,用磷酸盐缓冲液重悬菌体,沉淀后置冰盒超声破碎,4 ℃、13 684×g离心15 min分离蛋白上清和沉淀,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析,确定His-CdtA重组蛋白的最佳表达条件。
1.2.5 His-CdtA重组蛋白的纯化及鉴定 用最佳诱导条件大量诱导His-CdtA蛋白表达,利用镍柱亲和层析法,通过调节A液(磷酸盐缓冲液)和B液(500 mmol/L咪唑)比例,使咪唑浓度分别为10、50、100、200和300 mmol/L,探索His-CdtA重组蛋白的最佳纯化条件,SDS-PAGE验证表达及纯化结果。同时设立相同诱导条件下的pET-28b空载体表达蛋白阴性对照。
1.2.6 动物免疫 将纯化后的His-CdtA重组蛋白按100 μg/只给予6周龄的雌性Balb/c小鼠右后腿肌注,同时设立磷酸盐缓冲液阴性对照。分别在第1、7和14天进行小鼠免疫,首次免疫将His-CdtA重组蛋白与弗氏完全佐剂1∶1混匀,第2、3次免疫将His-CdtA重组蛋白与弗氏不完全佐剂1∶1混匀。最后1次免疫10 d后收集小鼠血清,分析抗CdtA抗体滴度。
1.2.7 His-CdtA重组蛋白的抗原性分析 将实验室保存的CdtA(C129-1392)重组蛋白按2 ng/μL包被96孔板,4 ℃包被过夜,分别加入100 μL按1∶102、1∶:103、1∶104和1∶105稀释的小鼠血清,做2个复孔,37 ℃孵育2 h后洗5次,加入100 μL按1∶104稀释的羊抗鼠IgG二抗-辣根过氧化物酶(horse radish peroxidase,HRP),37℃孵育1 h后洗5次,加入四甲基联苯胺显色液避光15 min后加入终止液,测450 nm处的A值。
1.2.8 His-CdtA重组蛋白的免疫印迹法鉴定 纯化后的His-CdtA重组蛋白经SDS-PAGE后经电转膜至PVDF膜,3%牛血清白蛋白封闭1 h,三羟甲基氨基甲烷-吐温缓冲液洗3次,以加入1∶500稀释的免疫后的小鼠血清为一抗,4 ℃振荡孵育过夜后洗3次,加入1∶104羊抗鼠IgG二抗-HRP,室温振荡孵育1 h后洗4次,加入混匀后的ECL工作液,在ECL呈像仪上曝光并保存结果。
2 结果
2.1 PCR扩增cdtA全长序列
将以RT 027型艰难梭菌菌株为模板的PCR产物经1%琼脂糖凝胶电泳后,在1 000~1 500 bp处有1条特异条带,序列大小与cdtA理论值1 392 bp相符。见图1。

图1 cdtA PCR产物的琼脂糖凝胶电泳图
2.2 菌液PCR鉴定pET-28b-cdtA扩增产物
随机挑取23个白色单菌落行菌液PCR,结果见图2。由于通用引物T7、T7ter包括载体自身片段约200 bp,故PCR扩增产物会比cdtA的理论值大,位于1 500 bp左右。选取14号菌为代表,测序后经Blast比对与GenBank上的艰难梭菌CdtA的序列100%匹配(登录号为HQ639678.1)。

图2 菌液PCR鉴定pET-28b-cdtA的琼脂糖凝胶电泳图
2.3 His-CdtA重组蛋白的最佳表达条件
当培养物A600达到0.6时,调节摇床温度为37 ℃,IPTG浓度为0,160 r/min 振荡培养12 h,此条件下的上清液在相对分子质量45 000上方有较多的蛋白表达,且根据迁移率计算该蛋白大小与His-CdtA重组蛋白的理论相对分子质量53 000相符(图3、图4),而阴性对照(泳道9)不表达(图3)。
2.4 His-CdtA重组蛋白的纯化
收集各浓度咪唑洗脱液行SDS-PAGE(图5)。上样穿透液(泳道2)目的蛋白明显减少,说明His-Cdt A重组蛋白已成功与镍柱结合,当洗脱液咪唑浓度为10 m mol/L时能有效洗脱杂蛋白(泳道3),直到浓度达到200 mmol/L时才能成功洗下目的蛋白(泳道6)。最终确定本研究His-CdtA重组蛋白的最佳纯化条件为:咪唑浓度为10 mmol/L时洗脱杂蛋白,咪唑浓度为200 mmol/L时完全洗脱His-CdtA目的蛋白。另外,pET-28b自身携带His标签,用相同条件表达纯化后虽然也有低浓度的蛋白被纯化,但与His-CdtA目的蛋白大小不一致(图4),进一步验证His-CdtA重组蛋白非载体自身表达蛋白。
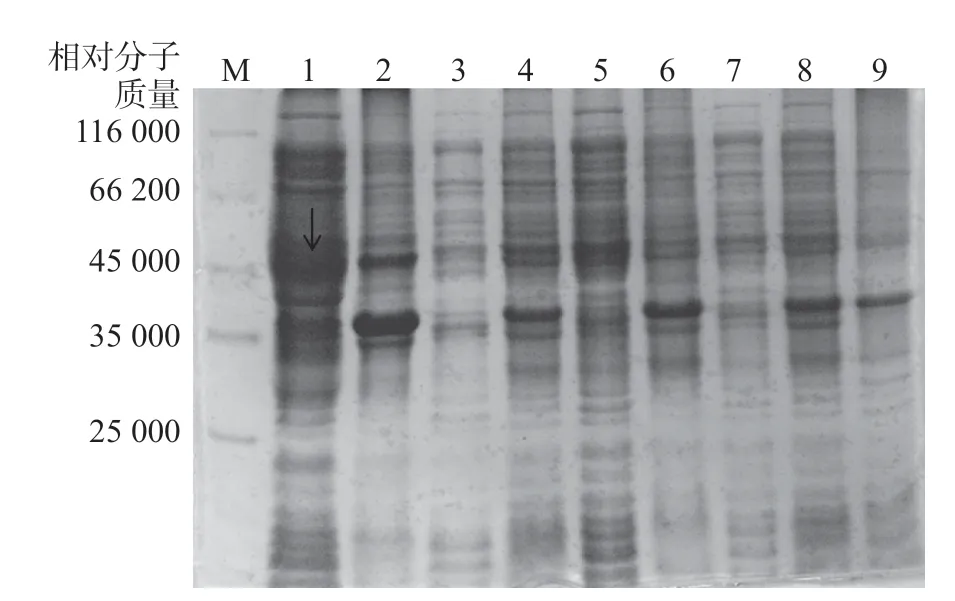
图3 诱导pET-28b-CdtA蛋白表达产物的SDS-PAGE图
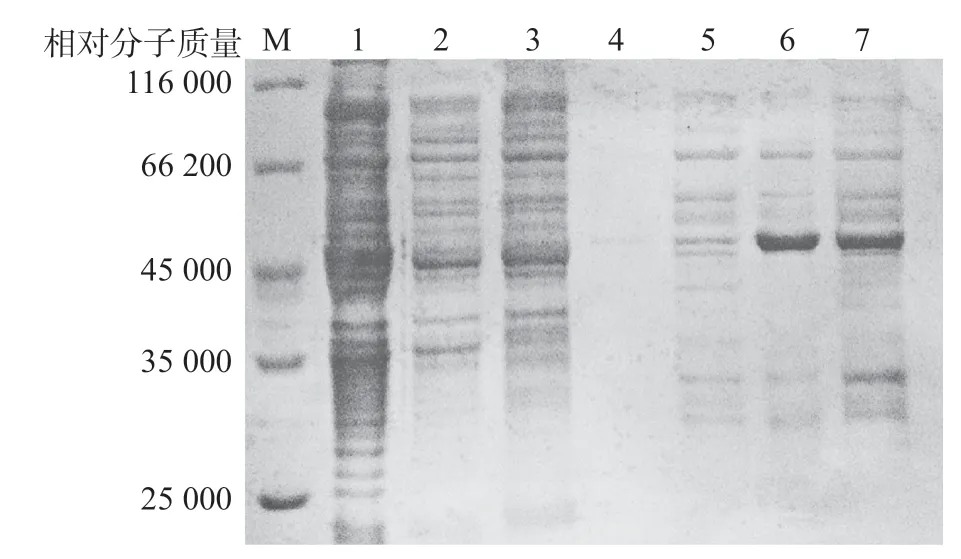
图4 镍柱亲和层析纯化产物的SDS-PAGE图
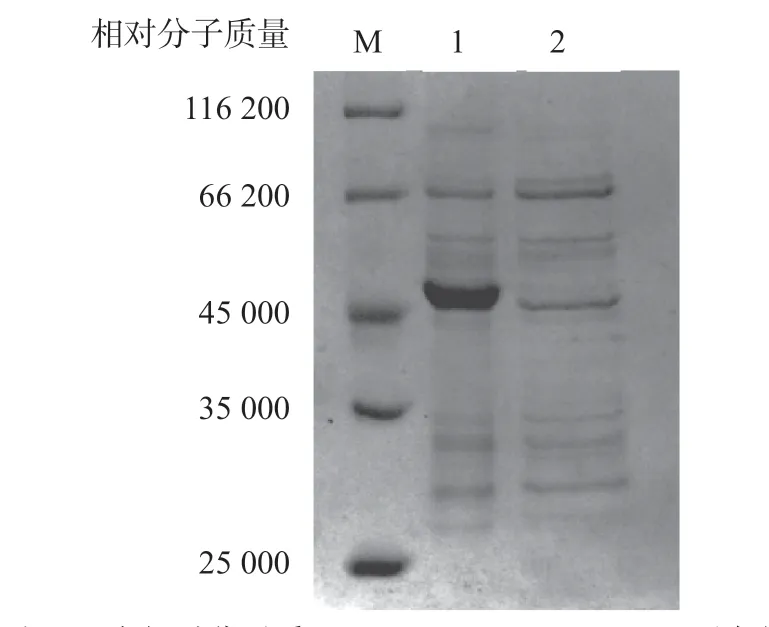
图5 重组菌与空白载体的表达蛋白纯化产物的SDS-PAGE图
2.5 His-CdtA重组蛋白的抗原性分析
免疫3次后的小鼠血清抗CdtA抗体效价达到1∶104。见表1。
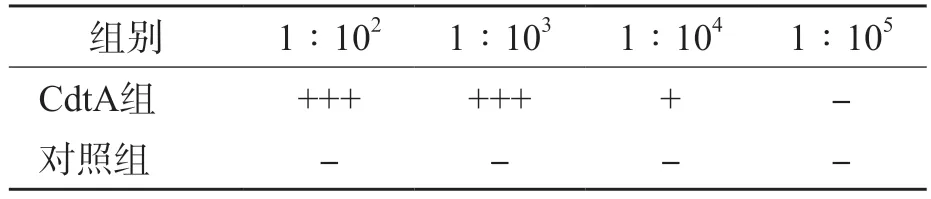
表1 His-CdtA重组蛋白的抗体滴度分析
2.6 His-CdtA重组蛋白的Western blot鉴定
免疫后的小鼠血清与纯化后的His-CdtA重组蛋白可特异性结合,而对应大小的pET-28b表达蛋白无此现象。见图6。
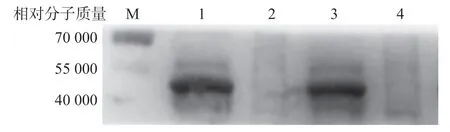
图6 His-CdtA重组蛋白的Western Blot鉴定结果
3 讨论
艰难梭菌可引起艰难梭菌相关性感染(Clostridium difficile-associated diarrhea,CDAD),是目前唯一能引起院内感染的革兰阳性厌氧芽孢梭菌,约25%的抗菌药物相关性腹泻、75%的抗菌药物相关性肠炎和约100%的假膜性肠炎均由此菌引起[12]。我国曾有1例老年患者感染RT 027型艰难梭菌导致休克的报道[13],2018年的1篇文献也报道了在分离自我国老年人群的116株艰难梭菌中发现1株RT 027型和6株RT 078型高产毒艰难梭菌[14],提示老年人群中存在高产毒艰难梭菌暴发感染的潜在风险,与非高产毒艰难梭菌比较,更严重、更易复发且预后差,但目前我国对艰难梭菌Cdt的研究非常少,更缺少可用的疫苗,故针对高产毒艰难梭菌菌株毒素抗原性的研究和疫苗的研制对预防艰难梭菌暴发感染十分重要。本研究成功构建了CdtA的全长基因(1 392 bp)原核表达载体pET-28b-CdtA,重组序列测序结果与GenBank中的CdtA序列(登录号为HQ639678.1)100%符合。实验中经诱导产生的可溶性His-CdtA重组目的蛋白通过镍柱亲和层析的方法纯化产物的相对分子质量为45 000~66 200,根据迁移率计算≈CdtA理论值(53 000),纯化的His-CdtA免疫小鼠后产生了高效价的抗CdtA抗体,经免疫印迹法鉴定,能与免疫后的小鼠血清特异结合,这些研究为后续相关蛋白单克隆抗体的制备和疫苗的研制奠定了基础。由于目前尚无商品化的艰难梭菌CdtA标准蛋白,本研究利用实验室保存的、以能产Cdt的标准菌株艰难梭菌(ATCCBAA-1870)为PCR模板构建pET-28b-CdtA(C129-1392)质粒,以表达并纯化的重组CdtA(C129-1392)作为标准蛋白包被96孔板,结果显示,小鼠产生的抗CdtA抗体的特异性和编码包被96孔板的本实验室保存的重组蛋白CdtA(C129-1392)的DNA来自产Cdt的标准菌株艰难梭菌(ATCC-BAA-1870),与本研究诱导制备的His-CdtA的DNA模板(加拿大圭尔夫大学Scott博士惠赠)的RT 027型艰难梭菌菌株的DNA来自不同菌株,二者虽多肽链片段长度有差异,但CdtA(C129-1392)与His-CdtA含有相同的抗原表位。本研究结果显示,本实验室保存的重组蛋白CdtA(C129-1392)能与实验中小鼠产生的抗CdtA抗体结合,间接验证了本研究诱导表达的CdtA重组蛋白His-CdtA的抗原特异性。由于目前尚无商品化的抗CdtA特异性一抗,Western blot实验结果显示,小鼠免疫后的血清与实验纯化的His-CdtA重组蛋白可特异性结合,进一步说明本研究的蛋白纯化产物为艰难梭菌CdtA。后续可考虑用His-CdtA刺激小鼠,制备相应的单克隆抗体,为Cdt的实验室检测及疫苗研制奠定基础。
