不同种类肉肌浆蛋白的油-水界面性质
2020-03-11杜菲菲吴长玲方艾虎杨宗韫李鹏鹏李典昭
杜菲菲,吴长玲,方艾虎,张 莉,李 震,杨宗韫,周 楠,李鹏鹏,李典昭,王 鹏*
(南京农业大学 肉品加工与质量控制教育部重点实验室,江苏 南京 210095)
随着全球人口的持续增加,人们对食品的需求量也在不断上升,使开发新的食物原料成为研究热点[1]。而我国的供给侧改革对食品质量提出了更高的要求,其目的是满足消费者对食品的更高感官属性及营养功能性。从以上两方面来看,开发新的食物原料并研究其加工特性势在必行。在肉糜生产加工过程中为去除肉腥味以及提高肌原纤维蛋白的凝胶性质,有时会去除肌浆蛋白,造成大量的浪费[2]。而畜禽肉在冷藏期间的汁液流失,也含有大量的肌浆蛋白[3]。肌浆蛋白是肌肉中含量较高的蛋白质,占蛋白总量的30%~35%[4]。在肌肉组织中,肌浆蛋白是天然存在的水溶性蛋白质并且溶于低离子强度的缓冲液中[5],主要是由肌溶蛋白、肌红蛋白和肌酶蛋白组成[6]。以往对于肌浆蛋白的研究多集中在其对于凝胶类肉制品的影响上[2,7-8]。
近年来,科研人员进一步挖掘肌浆蛋白的功能性,发现肌浆蛋白作为食品原料具有潜在应用价值,Sahin等[4]研究了电纺鱼肌浆蛋白纳米纤维的制备与表征,Yongsawatdigul等[9]研究表明含有40 单元转谷氨酰胺酶活力的罗非鱼肌浆蛋白能促进其肌动球蛋白的交联,交联程度随含量的增加而增加,同时促进肌动球蛋白重链和肌钙蛋白交联,但对肌动蛋白和原肌球蛋白无影响,肌浆蛋白可作为潜在的增强蜥蜴鱼肉酱凝胶强度的蛋白添加剂[10]。但针对不同种类动物肌浆蛋白界面性质的深入研究十分匮乏。本实验对猪肉、鸡肉和鱼肉肌浆蛋白的油-水界面性质进行了研究,并探究3 种蛋白的结构不同与功能差异的相关性,以期给相关研究人员提供借鉴。
1 材料与方法
1.1 材料与试剂
金龙鱼一级大豆油购于南京苏果超市。根据所占胴体的比例确定原料肉,猪肉取猪背最长肌,鱼肉选取鱼骨较少的鲶鱼,鸡肉选择鸡大胸,以上3 种肉均是苏果超市出售的新鲜产品。
25 mmol/L的磷酸盐缓冲液(25 mmol/L K2HPO4,25 mmol/L KH2PO4)、1-苯胺基萘-8-磺酸(8-anilino-1-naphthalenesulfonic acid,ANS)、双缩脲、牛血清白蛋白、5,5’-二硫代双-2-硝基苯甲酸(5,5’-dithiobis-(2-nitrobenzoic acid),DTNB)、磺基水杨酸 国药集团化学试剂有限公司;盐酸胍、十二烷基硫酸钠(sodium dodecyl sulfate,SDS) 阿拉丁控股集团有限公司。
1.2 仪器与设备
Avanti J-E离心机 美国Beckman Coulter公司;Ultra Turrax T-25 Basic高速匀浆机、C-MAG HS7磁力搅拌器 德国I K A公司;差示扫描量热仪美国PerkinElmer股份有限公司;L-8900日立全自动氨基酸分析仪 中国天美科学仪器有限公司;多功能酶标仪美国MD公司;Zeta电位仪 美国马尔文公司;界面流变仪 法国Teclis公司;Alpha 2-4 LSC plus冻干机德国Christ公司。
1.3 方法
1.3.1 不同肉类肌浆蛋白的制备
参照Liu Jiao等[11]的方法并加以修改。将肉块先用去离子水冲洗干净后,用手术刀切成小块,而后在绞肉机中绞碎,碎肉加入4 倍体积25 mmol/L的磷酸盐缓冲液(pH 7.2),再用组织破碎机在10 000 r/min匀浆10 s;匀浆液在4 ℃、13 000hg离心20 min,上清液用三层纱布过滤,滤液即为肌浆蛋白。
1.3.2 氨基酸组成测定
吸取约8 mL肌浆蛋白样品于离心试管中,3 000 r/min离心5 min(离心达到固液分离的目的),吸取离心后上清液1 mL,加入1 mL 2%的磺基水杨酸溶液,经涡旋仪振荡混匀后,静置10 min;在4 500 r/min离心20 min,离心后的上清液过0.45 µm的滤膜后装入2 mL的液相瓶中。
设置氨基酸自动分析仪的检测波长为440 nm和570 nm,设置反应柱温度35 ℃、流速0.400 mL/min、柱后衍生试剂流速0.350 mL/min。准确吸取混合氨基酸标准溶液,用pH 7.2的缓冲液稀释到5 mL,此标准溶液稀释浓度为5.00 nmol/50 µL,作为上机测定用的氨基酸标准溶液[12]。
1.3.3 理化指标测定
1.3.3.1 蛋白质溶解度
利用牛血清白蛋白作标准曲线,用双缩脲法在540 nm波长处测定蛋白质吸光度,测定结果为每毫升溶液含蛋白质的质量,即mg/mL。
1.3.3.2 粒径
采用配备4 mW氦-氖激光的Zetasizer Nano ZS 90,基于动态光散射(dynamic light scattering,DLS)原理进行分析[13],将3 种肌浆蛋白的质量分数利用25 mmol/L的磷酸盐缓冲液稀释到0.1%,每个样品重复测量3 次,取平均值。
1.3.3.3 蛋白质电位
方法同1.3.3.2节。
1.3.3.4 表面疏水性
参照Creamer[14]和常海霞[15]等的方法并加以修改。利用疏水性探针结合法进行测定,ANS作为荧光探针表征疏水性的强弱。将肌浆蛋白质量浓度稀释到1 mg/mL,并取4 mL至离心管,再加入20 µL 15 mmol/L的ANS(pH 7.0)溶液,利用涡旋振动仪充分振荡,在25 ℃(避光)静置20 min,再将样品滴入黑色96 孔板中,设置酶标仪的激发波长375 nm,发射波长410~570 nm,测定其荧光强度。
1.3.3.5 活性巯基含量
参照Ellman[16]和Guo Xiaoya[17]等的方法并加以修改。利用磷酸盐缓冲液将3 种肌浆蛋白的质量浓度统一稀释到1 mg/mL,取5 mL置于离心管中,加入20 µL DTNB,在涡旋仪上充分振荡后于室温(25 ℃)孵育1 h,将孵育完成后的样品滴入透明的96 孔板中,在412 nm波长处测定吸光度。巯基含量按下式计算:

式中:C0为巯基含量/(mol/g);A为412 nm波长处的吸光度;ε为摩尔消光系数(13 600 L/(molgcm));D为稀释倍数;ρ为蛋白质量浓度/(mg/mL)。
1.3.3.6 差示扫描量热法
参考Vieira等[18]方法并加以修改。差示扫描量热法是了解生物系统在原生状态下热活动的有利工具[19]。将3 种肉的肌浆蛋白溶液置于冻干机中冻干,然后分别称取样品15 mg,密封在铝盒中并放入样品池,以空盒作参比,保护气为氮气,升温速率8 ℃/min,温度范围20~110 ℃。
1.3.4 蛋白质构象稳定性分析
参照Kristinsson等[20]的方法并加以修改。利用6.0 mol/L的盐酸胍溶液将3 种肌浆蛋白质量浓度调到1 mg/mL后混合均匀,于25 ℃(室温)测定样品不同时间点的吸光度,测定范围为200~310 nm。由于构象变化速率可以更直观地反映蛋白质构象柔顺性,构象变化速率按下式计算:

1.3.5 肌浆蛋白界面行为及乳化性质分析
1.3.5.1 蛋白界面吸附动力学
参考Gao Zhiming等[21]方法,取大豆油500 mL,加入15 mL的Florisil吸附剂,搅拌30 min后,在5 000hg离心20 min;取出离心后的上清液继续加入吸附剂,重复上述操作3 次即得到纯化后的大豆油。利用界面流变仪测定纯水的动态界面张力,直到30 min内界面张力值下降不超过0.5 mN/m即满足要求。纯化后的大豆油密度为0.917 5 g/cm3,纯水与纯化后大豆油的界面张力为(26.5f0.5)mN/m。
实验过程中通过分析肌浆蛋白的界面张力(π)随着吸附时间的变化表征肌浆蛋白的界面吸附行为。测定在室温下进行,U形针浸入样品槽,样品槽放入25 mL相应的肌浆蛋白溶液(0.2 mg/mL),并通过马达控制形成10 µL大小的油滴,测定时间为10 800 s。测试过程中,外界不应有较大的振动,以免对测定造成较大的干扰。
1.3.5.2 肌浆蛋白乳化活性和乳化稳定性
参照李伟伟[22]的方法并加以修改。
乳化活性:将3 种肌肉的肌浆蛋白质量浓度稀释到1 mg/mL,然后加入30%体积的大豆油,在8 000 r/min转速下剪切2 min后,从底部取20 µL的乳化液置于50 mL离心管中,再加入1%的SDS溶液稀释100 倍,利用涡旋仪振动5 s后,在500 nm波长处测吸光度。
乳化稳定性:将上述制得的乳液在4 ℃静置10 min后,从底部取20 µL乳化液重复上述操作,在500 nm波长处测定吸光度。乳化活性指数和乳化稳定性的计算公式如下:

式中:T为浊度,T=2.303hA0;A0为500 nm波长处的吸光度;A10min为样品静置10 min时在500 nm波长处的吸光度,DF为稀释倍数;ρ为蛋白质质量浓度/(g/mL);φ为光路径1 cm;θ为油所占比例0.2。
1.4 数据处理
采用4 次重复实验的平均值利用SPSS 22.0对实验数据进行单因素方差分析,并用Origin 8.0作图。
2 结果与分析
2.1 3 种肉肌浆蛋白的氨基酸组成分析

表1 不同种类肉中肌浆蛋白的氨基酸含量Table 1 Amino acid contents of sarcoplasmic proteins in meat from different animal species
氨基酸的组成及排列顺序影响蛋白质的营养特性、生化活性以及加工特性。有些氨基酸有较强的疏水性,因此这些氨基酸很大程度上会增加蛋白质的表面疏水性,进而对蛋白质的功能特性产生影响。因此,蛋白质在油-水界面稳定性的诸多影响因素中,蛋白质暴露于界面的氨基酸组成不容忽视,特别是苯丙氨酸和酪氨酸这些疏水性极强的氨基酸,对保持蛋白质三级结构也起着重要作用。从表1可以发现,鸡肉肌浆蛋白中这两种氨基酸含量最高,且与其他两种肌肉肌浆蛋白间有显著性差异(P<0.05)。
2.2 肌浆蛋白的基本理化指标测定结果分析

表2 不同种类肉肌浆蛋白理化指标的测定结果Table 2 Physicochemical indexes of sarcoplasmic proteins in meat from different animal species
蛋白质的表面性质涉及到蛋白质在极性不同的两相之间产生的作用,对于研究蛋白质的性质等具有重要意义。影响蛋白质在油-水界面上吸附的因素有很多,主要是蛋白质的氨基酸组成及其序列分布、形状、分子粒径、构象、表面疏水性、电荷大小等。溶解度是反映蛋白质溶液状态和聚集程度的重要参数,肌肉蛋白质的溶解度是指在特定的提取条件下溶解于水溶液中的原蛋白质百分比,它是溶质(蛋白质)和溶剂(水)之间平衡的表现[23],如表2所示,对于相同条件提取的蛋白,鱼肉肌浆蛋白溶解度显著最大达到(16.74f0.39)mg/mL(P<0.05),而猪肉肌浆蛋白与鸡肉肌浆蛋白均在9.5~10.6 mg/mL之间,后面的二者间溶解度无显著差异(P>0.05)。从粒径的角度看,大粒径极大可能在空间产生位阻,进而削弱表面疏水性的提高带来的扩散速率增加。鸡肉肌浆蛋白的粒径为(231.7f7.74)nm,相对稍大于其他两种肌浆蛋白,但是整体无显著差异(P>0.05),这一现象与吸附动力学具有理论关联性。蛋白质电位是衡量体系稳定性的重要指标[24],因为它不仅反映颗粒所带电荷的大小,而且还能表征颗粒间相互作用的强弱,Zeta电位越高,即所带电荷数越多,体系就越稳定。3 种肉肌浆蛋白Zeta电位分别为(-12.65f1.39)、(-13.64f1.57)、(-15.64f1.99)mV左右,其中鱼肉肌浆蛋白的Zeta电位相对于其他两种蛋白显著较高(P<0.05),Zeta电位不仅影响着肌浆蛋白质分子间的相互作用,而且在蛋白质的凝胶及乳化方面也有着显著性的影响[25]。不同肉类肌浆蛋白间表面疏水性的差异可能是由于其氨基酸组成、亚基组成和蛋白构象不同所导致,由于蛋白质中苯丙氨酸和酪氨酸的疏水性极强,而由表1可知,鸡肉肌浆蛋白中这两种氨基酸含量都要高于其他两种肌浆蛋白,所以这也很好地解释了鸡肉肌浆蛋白的表面疏水性在3 种肌浆蛋白中最大。表面疏水性同时又与其他指标有着密不可分的关系,例如热变性温度,因为适当的热处理会造成肌浆蛋白内部疏水基团暴露在蛋白质表面,从而使蛋白质的表面疏水性增强,这样就可以改善蛋白质的乳化活性和乳化稳定性。3 种肌浆蛋白热变性温度都较低,在40~55 ℃之间,其中鱼肉肌浆蛋白变性温度最高而猪肉肌浆蛋白最低,且猪肉肌浆蛋白热变性温度与鸡肉和鱼肉间差异显著(P<0.05),而鸡肉与鱼肉肌浆蛋白间的变性温度无显著性差异。所以在肌浆蛋白的研究中,如何根据不同蛋白的变性温度差异,准确控制热处理的强度至关重要,本研究可给蛋白变性凝聚组装类的研究提供基础数据的参考。
2.3 蛋白质构象稳定性的结构分析

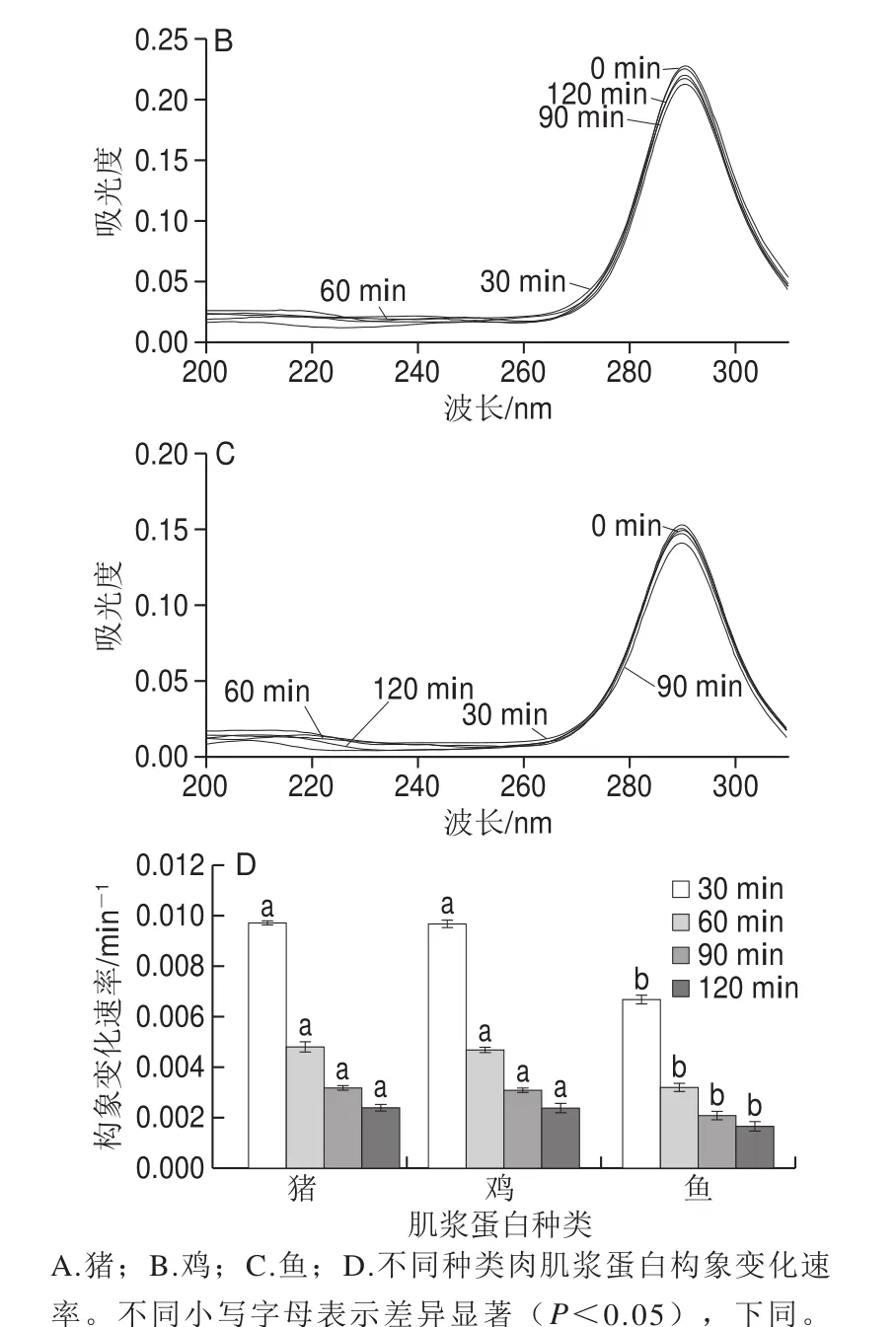
图1 不同种类肉肌浆蛋白在不同时间点吸光度及构象速率的变化Fig. 1 UV absorption spectra and conformational change rates of sarcoplasmic proteins in meat from different animal species at different time points
对于阐述蛋白质表面疏水性与其乳化活性的关系方面已有大量研究支撑,但是在蛋白质构象特性与乳化活性关系方面却鲜有研究,这是因为蛋白质的结构会随着周围环境的变化而变化,导致蛋白质构象特性对其油-水界面性质的表征有很大难度。本实验主要利用盐酸胍诱导蛋白质变性,从而探究在不同处理时间下蛋白质构象的动态变化,盐酸胍的加入能破坏蛋白质的氢键、疏水键等,进而能破坏蛋白质的高级结构。而蛋白质在油-水界面的构象展开也可以被视为一种类似于部分“变性”的行为,因此盐酸胍诱导蛋白质结构展开是模拟蛋白质在油-水界面构象变化的良好手段,从而体现在油-水界面上的构象柔顺性,以及完成空间定位的难易程度[26],所以本研究对于揭示不同蛋白乳化特性差异有很重要的作用。盐酸胍诱导肌浆蛋白变性的荧光光谱如图1A~C所示,3 种肌浆蛋白都在290 nm波长处达到最大吸光度,由于酪氨酸主要对250~290 nm波长范围内具有吸收。由肌浆蛋白氨基酸组成可知,鸡肉肌浆蛋白的酪氨酸含量最高,这也解释了在第0分钟时鸡肉肌浆蛋白的吸光度相比较而言达到最大。而且猪肉和鱼肉肌浆蛋白在第120分钟时达到最大吸光度,这说明肌浆蛋白三级结构改变时,构象发生重排且构象展开,而鸡肉肌浆蛋白在第120分钟时吸光度有所下降,说明鸡肉肌浆蛋白结构改变较快。由图1D可知,随着时间的延长,构象变化速率越来越小,并且在相同时间下,猪肉肌浆蛋白与鸡肉肌浆蛋白的峰值变化速率无显著差异(P>0.05),而鱼肉肌浆蛋白峰值变化速率相比较而言显著较慢(P<0.05)。这一结果表明,和鱼肉肌浆蛋白相比,猪肉和鸡肉的肌浆蛋白构象柔顺性更好。
2.4 肌浆蛋白界面吸附动力学分析
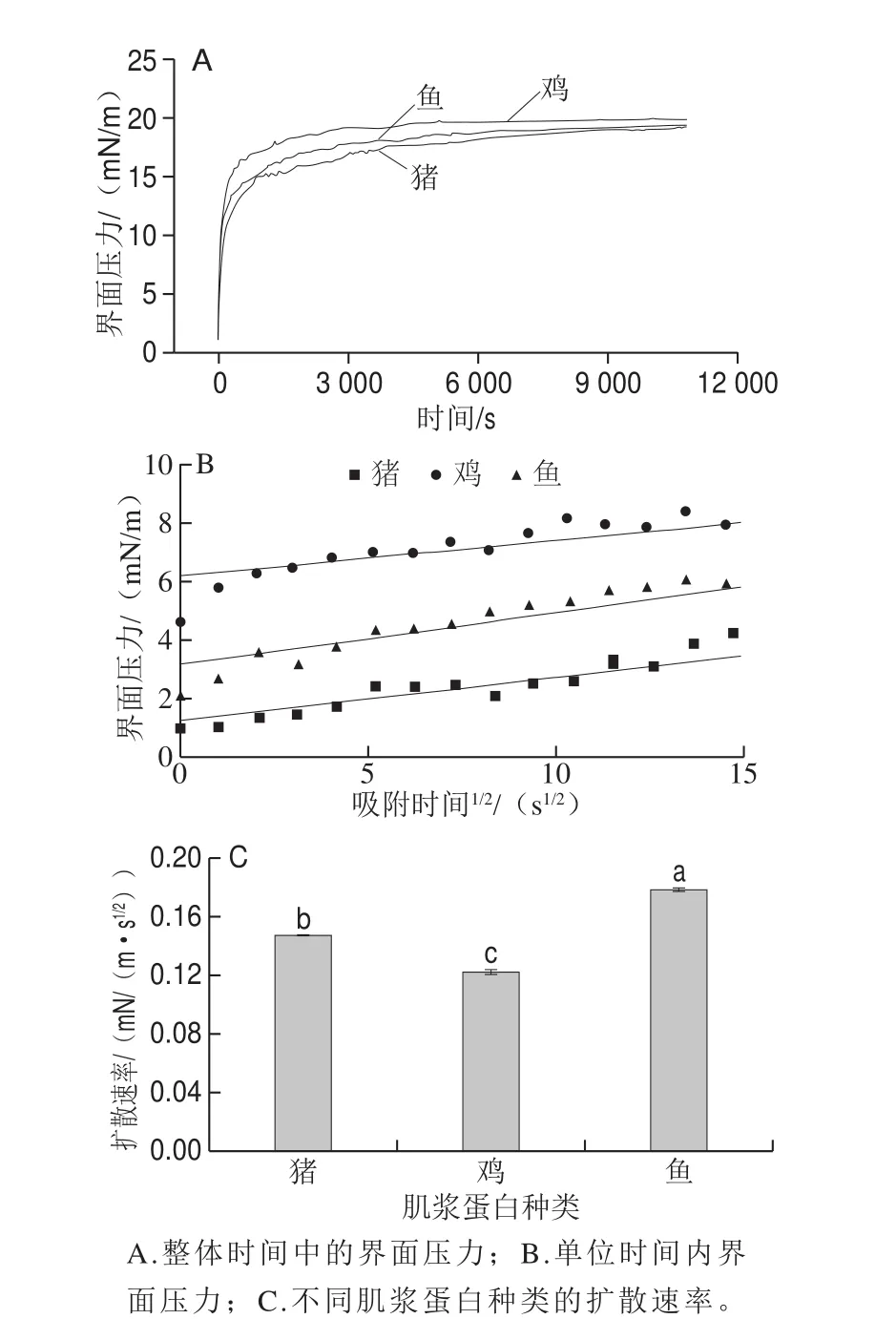
图2 不同种类肉肌浆蛋白界面压力与吸附速率的变化Fig. 2 Changes in interfacial pressure and adsorption rate of sarcoplasmic proteins in meat from different animal species
界面流变学提供了流体界面上分子吸附的重要信息[27]。通过测定界面压力可以表征O/W界面的性质和表面活性物质的吸附和解吸附动力学性质,一般来说,油水界面蛋白质的吸附动力学过程包括以下几个重要步骤:首先蛋白质在界面上扩散形成一个次级界面区域;然后到达界面的蛋白质分子部分展开,界面张力下降;最后吸附的蛋白质分子在油-水界面发生结构重排[28]。
越来越多的研究表明[29-30],界面流变性质极大影响着蛋白质的乳化性质,且两者关系紧密。在利用蛋白质制备乳液的过程中,蛋白质会吸附到油滴表面使得油水界面张力降低,所以张力是影响乳状液稳定性的一个主要因素。乳状液的形成必然使体系面积大大增加,也就是对体系要做功,从而增加了体系的界面能,这成为体系不稳定的来源,因此深入研究蛋白质的油-水界面性质对于揭示分散系的稳定机理有重要意义。本研究中,动态界面张力通过界面压力随吸附时间的变化表示,表面张力是由缓冲液的界面张力减去蛋白的界面张力所得。用直线的斜率表示扩散速率,柱状图即为吸附动力学特征参数。从图2可以明显看出,随着吸附时间的延长,界面压力增大,表明蛋白质逐渐吸附到油-水界面,并且在最初的5 min内3 种肌浆蛋白的界面压力快速增加。其中猪肉肌浆蛋白的界面压力在初期增长显著高于其他两种肌浆蛋白,这一现象的发生可能是由于猪肉肌浆蛋白的疏水性高于其他两者,根据Li Lingyun等[31]的研究可知表面疏水性是影响蛋白质对界面油侧吸附的关键因素之一。此后3 种肌浆蛋白界面压力值的增长趋缓,且鸡肉肌浆蛋白的界面压力始终大于其他两者,而猪肉肌浆蛋白界面压力最低。当吸附时间达到180 min后,界面压力趋于稳定,表明吸附基本达到平衡。在界面压力π值增加的起始阶段,界面上蛋白质浓度较低,吸附动力学受扩散作用的影响,遵循变形后的Ward和Tordai扩散模型所描述的π随吸附时间的变化[32],如下式所示:
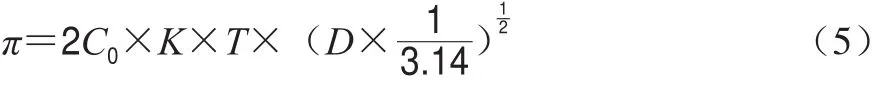
式中:C0为初始浓度;K为波兹曼常数;T为绝对温度;D为扩散系数。
由图2可知,不同种类肌浆蛋白的动力学扩散速率间存在差异性。蛋白质分子在界面上的扩散速率受其粒径大小的影响,而本实验结果更能解释这一现象。在前述理化指标中,粒径大小为鸡肉肌浆蛋白>猪肉肌浆蛋白>鱼肉肌浆蛋白,而在本部分扩散速率大小为鸡肉肌浆蛋白<猪肉肌浆蛋白<鱼肉肌浆蛋白。这可能是由于大粒径的肌浆蛋白产生的空间位阻在一定程度上削弱了表面疏水性提高带来的吸附速率增加。而且鱼肉肌浆蛋白的扩散速率较猪肉和鸡肉肌浆蛋白间显著较大(P<0.05),但鸡肉与猪肉肌浆蛋白之间的扩散速率却无显著性差异(P>0.05)。
2.5 肌浆蛋白乳化活性及乳化稳定性

图3 不同种类肉肌浆蛋白乳化活性及乳化稳定性的差异Fig. 3 Differences in emulsifying activity and emulsion stability of sarcoplasmic proteins in meat from different animal species
在前面的研究中得到,蛋白质的疏水性及粒径大小等会影响其油-水界面的性质,而蛋白质作为乳化剂其界面特性在它们形成和稳定乳液能力方面起着重要作用[33],进而影响蛋白质的乳化活性。油-水界面属于高能界面,未加乳化剂形成的乳化体系会很快崩溃,蛋白质是两性物质,既有疏水基团也有亲水基团,这样才使得蛋白质具有独特的乳化性能,且蛋白质表面疏水性越大,蛋白质所制得的乳液稳定性也会增强[34]。对于不同肉类同种蛋白,表面特性和构象特性的差异会引起乳化活性和乳化稳定性的不同,如图3所示。鸡肉肌浆蛋白乳化活性最高,猪肉肌浆蛋白乳化活性稍低于鸡肉但无显著差异性(P>0.05),但是鱼肉肌浆蛋白乳化活性显著低于猪肉和鸡肉肌浆蛋白(P<0.05),这一结论与蛋白构象稳定性结果相符合。在盐酸胍诱导蛋白变性时,随时间的增加使其形成刚性结构,这正是导致鱼肉肌浆蛋白的峰值变化速率相比较猪肉和鸡肉而言差异性显著且其乳化活性较低的原因。鸡肉肌浆蛋白的稳定性相对于猪肉肌浆蛋白有所下降但差异性不显著,可能是因为鸡肉肌浆蛋白粒径相对最大,导致其形成乳液液滴变大,而一般来说乳液液滴越小越稳定[35],所以粒径可能是导致鸡肉肌浆蛋白稳定性变差的原因。而3 种肌浆蛋白中鱼肉肌浆蛋白的乳化稳定性显著最差(P<0.05),这可能与鱼肉肌浆蛋白的构象稳定性相对较差有关。
2.6 不同种类肉肌浆蛋白各指标的主成分分析
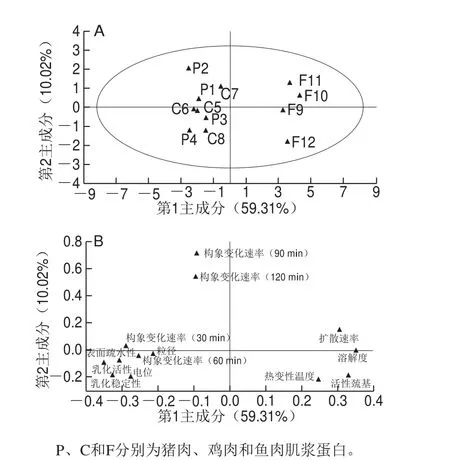
图4 不同种类肉肌浆蛋白主成分分析散点图(A)和载荷图(B)Fig. 4 Scatter plot (A) and loading plot (B) of principal components analysis for sarcoplasmic proteins in meat from different animal species
由于测量指标间存在线性相关性,单一的指标不能更清楚地反映出不同种类肉肌浆蛋白间的差异性,因此进一步进行主成分分析。由图4A可知,第1主成分占59.31%,第2主成分占10.02%,总共占比69.33%,其中样本F10、F11以及F12对第1主成分贡献率较大,且与第1主成分呈正相关,而C7样本对第2主成分贡献率较大且呈正相关。由图4B可知,肌浆蛋白的扩散速率、溶解度、乳化活性、乳化稳定性、表面疏水性、电位、构象在30 min及60 min时的变化速率对第1主成分贡献率较大,其中扩散速率和溶解度与第1主成分呈正相关,而除此以外的其他指标与第1主成分呈负相关。而蛋白的构象在90 min以及120 min时的变化速率对第2主成分的贡献率较大。图中的乳化活性、表面疏水性、乳化稳定性及电位间距离较近,说明这些指标间存在一定的相关性。综合前述的研究结论可知肌浆蛋白的乳化活性和乳化稳定性与其表面疏水性、电位和构象变化速率有较强的相关性,而扩散速率与溶解度间相关性较强。
3 结 论
本实验以猪肉、鸡肉及鱼肉的肌浆蛋白为研究对象,通过测定其理化性质、氨基酸组成、蛋白质构象、界面性质以及乳化性质进而深入了解其油-水界面的性质。研究结果表明:不同种类肉的肌浆蛋白其理化性质、氨基酸组成、蛋白质构象、界面性质以及乳化性质都有一定的差异性。
肌浆蛋白中氨基酸组成、粒径和变性温度等都显著影响蛋白疏水性,其中鸡肉肌浆蛋白中酪氨酸和苯丙氨酸含量较高以致疏水性高于其他两种肌浆蛋白,而3 种肉肌浆蛋白的粒径间无显著差异,但猪肉肌浆蛋白与鸡肉肌浆蛋白和鱼肉肌浆蛋白间有显著差异。
对于盐酸胍诱导的蛋白质构象稳定性,当肌浆蛋白三级结构改变时,构象发生重排且构象展开,鱼肉肌浆蛋白峰值变化速率相比较而言显著较慢,说明猪肉与和鸡肉的肌浆蛋白结构构象相对于鱼肉肌浆蛋白柔顺性更强。
肌浆蛋白的吸附动力学实验表明鱼肉肌浆蛋白的扩散速率较猪肉和鸡肉肌浆蛋白间显著较大,这可能是由于大粒径的肌浆蛋白产生的空间位阻在一定程度上削弱了表面疏水性的提高带来的吸附速率的增加,但鸡肉与猪肉肌浆蛋白之间的扩散速率却无显著性差异。
实验结果还表明,鸡肉肌浆蛋白乳化活性最高,鱼肉肌浆蛋白乳化活性显著低于猪肉和鸡肉肌浆蛋白,猪肉肌浆蛋白乳化活性稍低于鸡肉但无显著差异性,这可能是由于肌浆蛋白粒径及构象的不同造成的。但鱼肉肌浆蛋白的乳化稳定性显著最差。
本研究以分子间作用力及分子结构分析和界面流变研究作为有力工具,揭示肌浆蛋白纳米颗粒的界面吸附动力学规律。围绕这个中心可再进行深入研究,例如利用乳液替代食品中饱和脂肪的功能性食品等,利用肌浆蛋白纳米颗粒皮克林乳液调控脂肪消化,为功能性食品的开发提供理论依据。
