奇果菌素调控miR-646/ADAM17通路对白血病细胞增殖、凋亡的影响及机制
2020-03-03努尔阿米娜依明尼亚孜帕力旦艾沙阿依姆妮萨阿卜杜热合曼
努尔阿米娜·依明尼亚孜 帕力旦·艾沙 阿依姆妮萨·阿卜杜热合曼
急性淋巴细胞白血病属于血液系统性疾病,其主要特征是细胞恶性增殖、细胞凋亡受抑[1]。因而如何抑制白血病细胞增殖及促进细胞凋亡成为目前研究重点问题。研究表明奇果菌素(grifolin,Gri)可通过诱导人骨肉瘤细胞凋亡从而抑制骨肉瘤细胞生长[2]。奇果菌素还可抑制胃癌细胞增殖及侵袭,诱导细胞凋亡[3]。但奇果菌素对白血病细胞增殖、凋亡的影响尚未可知。研究表明微小RNA-646(microRNA-646,miR-646)在结直肠癌、胃癌、骨肉瘤组织及细胞中下调表达,并可抑制细胞增殖及迁移[4-6]。但miR-646对白血病细胞增殖、凋亡的影响尚未可知。TargetScan预测显示解聚素-金属蛋白酶17(a disintegrin and metalloprotease 17,ADAM17)可能是miR-646的靶基因,研究表明ADAM17在食管鳞状细胞癌组织和细胞中呈高表达,其高表达量与细胞侵袭及转移有关[7]。沉默ADAM17表达可抑制人甲状腺乳头状癌细胞增殖、迁移及侵袭[8]。但笔者发现miR-646与ADAM17的靶向关系,以及奇果菌素是否通过调控miR-646/ADAM17通路影响白血病细胞的增殖和凋亡尚未可知。本研究以白血病K562细胞为研究对象,深入探讨奇果菌素对白血病细胞的增殖和凋亡影响及其对miR-646/ADAM17通路的调控作用。
1 材料与方法
1.1 材料与试剂 奇果菌素购自上海赫澎生物科技有限公司;白血病细胞K562购自美国ATCC细胞库。RPMI 1640培养基购自上海源叶生物科技有限公司;甲基噻唑基四唑(methylthiazolyl tetrazolium,MTT)细胞增殖试剂盒购自上海齐一生物科技有限公司;细胞凋亡试剂盒购自北京庄盟国际生物基因科技有限公司;Trziol试剂、反转录与实时荧光定量PCR试剂盒与聚氰基丙烯酸正丁酯(bicinchonininc acid,BCA)蛋白浓度测定试剂盒均购自美国Thermo Fisher公司;磷酸盐缓冲液(phosphate buffered saline,PBS)、蛋白提取试剂盒购自上海联迈生物工程有限公司;miR-646 模拟物(mimics)、阴性对照(miR-NC)、miR-646抑制剂(anti-miR-646)、anti-miR-NC、ADAM17小干扰RNA(si-ADAM17)、乱序无意义阴性对照序列(si-NC)购自上海吉玛基因;Lipofectamine2000购自上海北诺生物科技有限公司;兔抗人细胞周期蛋白1(CyclinD1)、P21一抗购自美国Cell Signaling Technology公司;兔抗人ADAM17、 B淋巴细胞瘤-2 (B-cell lymphoma-2,Bcl-2)、B淋巴细胞瘤-2相关蛋白(Bcl-2-associated X protein,Bax)单克隆抗体购自美国Abcam公司;辣根过氧化物酶(HRP)标记的山羊抗兔IgG二抗购自武汉艾美捷科技有限公司。
1.2 方法
1.2.1 实验处理与分组:白血病K562细胞培养于RPMI 1640完全培养基(10%胎牛血清),放入37℃恒温培养箱进行培养,每24小时更换一次培养液,待细胞生长汇合至90%时进行传代培养。用10%胎牛血清的RPMI 1640培养液稀释奇果菌素,稀释浓度为25 μmol/L、50 μmol/L、75 μmol/L[9]。选取对数生长期白血病K562细胞,用终浓度为25 μmol/L、50 μmol/L、75 μmol/L的奇果菌素分别作用24 h,分别为Gri 25 μmol/L组、Gri 50 μmol/L组、Gri 75 μmol/L组,未经任何处理的K562细胞作为Con组。同时将miR-646 mimics(miR-646组)、miR-NC(miR-NC组)、si-ADAM17(si-ADAM17组)、si-NC(si-NC组)分别转染至K562细胞,各组细胞培养48 h。后续实验中将anti-miR-646、anti-miR-NC分别转染至K562细胞后,用终浓度为75 μmol/L的奇果菌素作用24 h。
1.2.2 细胞增殖能力检测:取对数生长期K562细胞,加入RPMI 1640培养液调整细胞密度为1×104个/ml,接种于96孔板(3×104个/孔),继续培养24 h,按照“1.2.1”分组处理,同时设置空白对照组,每组设置3个复孔,各组处理24 h时每孔加入20 μl MTT溶液(5 mg/ml),37℃恒温培养箱继续培养4 h,弃上清,每孔加入150 μl二甲基亚砜(DMSO),低速振荡10 min,应用酶标仪检测各孔吸光度值(A值),计算细胞抑制率=[1-(试验组A值-空白组A值)/(对照组A值-空白组A值)]×100%,实验重复3次。
1.2.3 细胞凋亡实验:取各组对数生长期K562细胞,(室温条件下)1 000 r/min转速离心6 min,弃上清,PBS清洗后加入500 μl结合缓冲液,加入5 μl Annexin V-FITC,充分混匀后加入5 μl PI,室温避光孵育10 min,应用流式细胞仪检测细胞凋亡率。
1.2.4 实时荧光定量聚合酶链反应(Quantitative Real-time PCR,qRT-PCR)检测细胞中miR-646、ADAM17 mRNA表达水平:取各组K562细胞,采用Trizol法提取细胞总RNA,应用nanodrop微量核酸测定仪测定RNA浓度,取2 ng RNA进行反转录反应,严格按照试剂盒说明书配置反应体系,经反转录得到cDNA,以cDNA为模板配置qRT-PCR反应体系,反应条件为95℃ 2 min(循环1次),95℃ 30 s,60℃30 s,72℃ 30 s(循环35次),严格按照试剂盒说明书进行操作,qRT-PCR反应所用仪器为ABI 7500型荧光定量PCR仪,反应结束后,miR-646以U6为内参,ADAM17以GAPDH为内参,采用2-ΔΔCt法计算miR-646、ADAM17 mRNA相对表达量。miR-646正向引物为5’-ATAGGCCGGCATAGACGCGTCTGGCT-3’,反向引物为:5’-AAAGATCCTTTATTAAGCTTAATGGGA-3’;ADAM17正向引物为5’-ATCAAACCCTTTCCTGCG-3’,反向引物为:5’-CAAACCCATCCTCGTCCA-3’。
1.2.5 蛋白免疫印迹法(Western blot)检测ADAM17、CyclinD1、Bcl-2、p21、Bax蛋白表达:收集各组K562细胞,参照蛋白提取试剂盒提取细胞总蛋白,严格按照BCA蛋白浓度测定试剂盒说明书测定蛋白浓度,蛋白高温变性,SDS-PAGE电泳的加样孔内加入变性蛋白(30 μg/孔),电泳结束后转移至PVDF膜,用5%脱脂牛奶封闭PVDF膜2 h,加入一抗(1∶1 000),摇床上4℃孵育过夜,TBST洗涤3次×10 min,加入二抗(1∶5 000),室温孵育1 h,TBST洗涤3次×10 min,滴加ECL显影,应用凝胶成像分析系统与ImageJ软件分析蛋白条带灰度值。
1.2.6 双荧光素酶报告基因检测:构建野生型荧光素酶报告载体WT-ADAM17与突变型荧光素酶报告载体MUT-ADAM17,WT-ADAM17、MUT-ADAM17分别与miR-646 mimics、miR-NC共转染至K562细胞,置于37℃恒温饱和湿度培养箱继续培养24 h,收集细胞后检测荧光素酶活性。
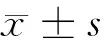
2 结果
2.1 奇果菌素对白血病K562细胞增殖、凋亡的影响 与Con组相比,Gri 25 μmol/L组、Gri 50 μmol/L组、Gri 75 μmol/L组白血病K562细胞抑制率显著升高(P<0.05),细胞凋亡率显著增加(P<0.05),CyclinD1、Bcl-2蛋白表达水平显著降低(P<0.05),p21、Bax蛋白表达水平显著升高(P<0.05),Gri 不同剂量组间比较差异具有统计学意义(P<0.05),且呈剂量依赖效应。见图1,表1。
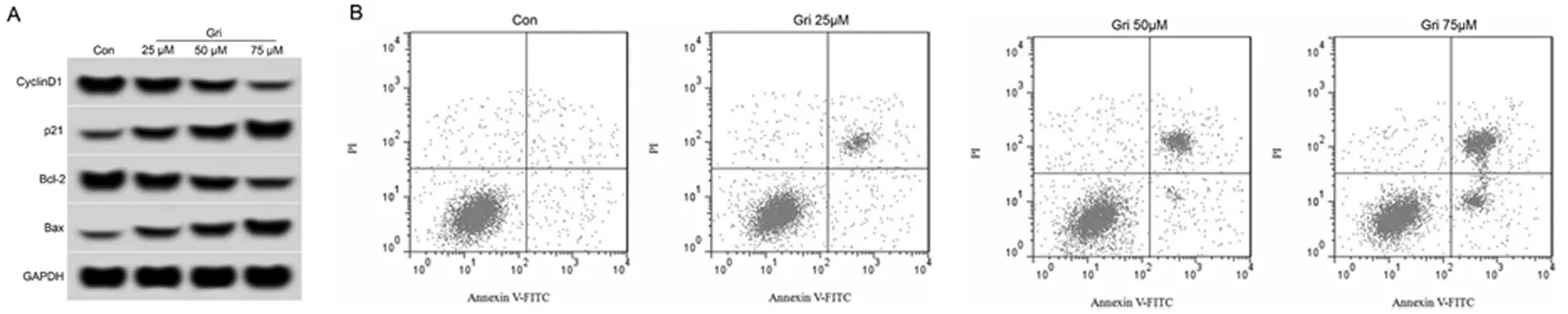
图1 奇果菌素对白血病K562细胞增殖、凋亡的影响;A 增殖、凋亡相关蛋白表达;B 细胞凋亡流式图
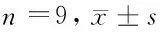
表1 奇果菌素对白血病K562细胞增殖、凋亡的影响
2.2 奇果菌素对白血病K562细胞中miR-646和ADAM17表达的影响 实验结果显示,相对于Con组,Gri 25 μmol/L组、Gri 50 μmol/L组、Gri 75 μmol/L组白血病K562细胞中miR-646的表达低水平显著升高(P<0.05),ADAM17的表达水平显著降低(P<0.05),奇果菌素不同剂量组间比较差异有统计学意义(P<0.05),且呈剂量依赖效应。见图2,表2。
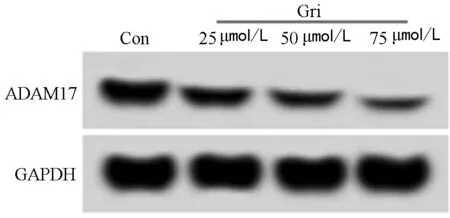
图2 ADAM17蛋白表达
表2 奇果菌素对白血病K562细胞中miR-646和ADAM17表达的影响
2.3 miR-646靶向调控ADAM17的表达 TargetScan预测显示ADAM17的3’UTR中含有与miR-646互补的核苷酸序列,见图3A。双荧光素酶报告实验结果显示,miR-646过表达可抑制含miR-646结合位点荧光报告载体的强度(P<0.05),而含突变结合位点的荧光报告载体的强度无显著性变化。Western blot实验结果显示,miR-646组白血病K562细胞中ADAM17的表达水平显著低于miR-NC组(P<0.05),anti-miR-646组白血病K562细胞中ADAM17的表达水平显著高于anti-miR-NC组(P<0.05)。见图3,表3、4。
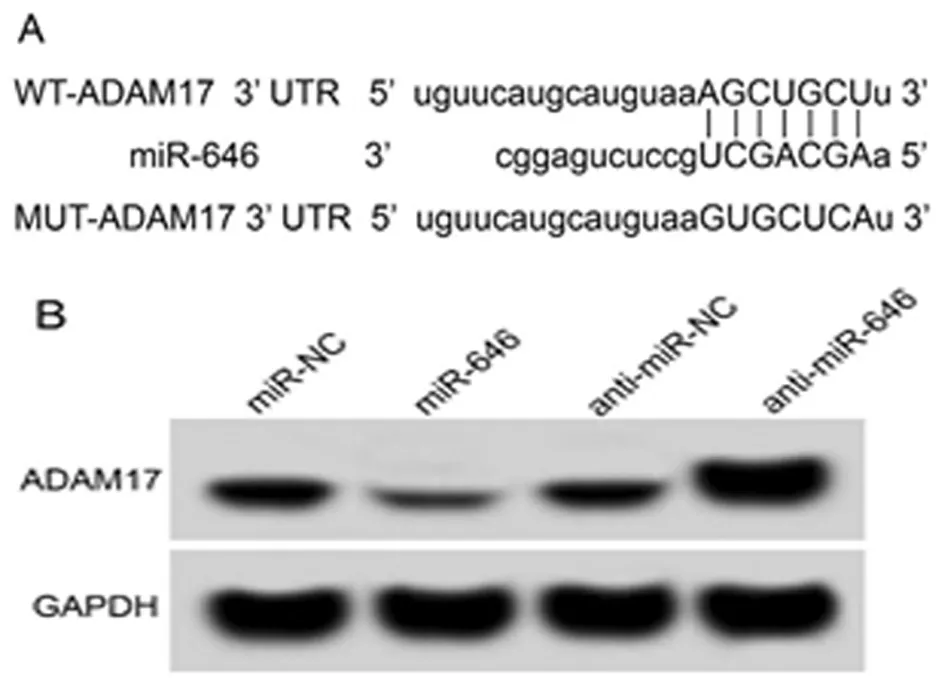
图3 miR-646靶向调控ADAM17的表达;A ADAM17的3’UTR中含有与miR-646互补的核苷酸序列;B ADAM17蛋白表达
表3 双荧光素酶报告实验
表4 miR-646调控ADAM17蛋白的表达
2.4 miR-646过表达对白血病K562细胞增殖、凋亡的影响 与miR-NC组相比,miR-646组白血病K562细胞抑制率与细胞凋亡率均显著升高(P<0.05),CyclinD1、Bcl-2蛋白的表达水平显著降低(P<0.05),p21、Bax蛋白的表达水平显著升高(P<0.05)。见表5,图4。
表5 miR-646过表达对血病K562细胞增殖、凋亡的影响

图4 增殖、凋亡相关蛋白表达
2.5 干扰ADAM17表达对白血病K562细胞增殖、凋亡的影响 相较于si-NC组,si-ADAM17组白血病K562细胞抑制率与细胞凋亡率均显著增加(P<0.05),CyclinD1、Bcl-2蛋白表达下调(P<0.05),p21、Bax蛋白表达上调(P<0.05)。见图5,表6。

图5 ADAM17和增殖、凋亡相关蛋白表达
表6 干扰ADAM17表达对血病K562细胞增殖、凋亡的影响
2.6 抑制miR-646表达逆转了奇果菌素(75 μmol/L)对白血病K562细胞增殖、凋亡的作用 相对于Gri+anti-miR-NC组,Gri+anti-miR-646组白血病K562细胞中ADAM17蛋白表达水平显著升高(P<0.05),细胞抑制率与细胞凋亡率均显著减低(P<0.05),CyclinD1、Bcl-2蛋白的表达水平显著升高(P<0.05),p21、Bax蛋白的表达水平显著降低(P<0.05)。见表7,图6。
表7 抑制miR-646表达逆转了奇果菌素对白血病K562细胞增殖、凋亡的作用
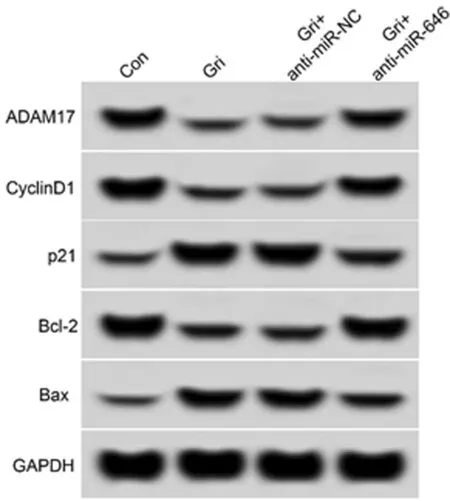
图6 ADAM17和增殖、凋亡相关蛋白表达
3 讨论
白血病发生发展过程涉及基因遗传、基因突变等多种因素,临床采用骨髓移植、干细胞移植等治疗白血病,但移植过程中极易引发炎症从而影响患者生存质量[10]。分子靶向治疗为白血病的治疗提供新方向,研究显示miRNA可通过调控下游靶基因表达从而参与白血病发生过程,miRNA可能作为靶向治疗白血病的潜在靶点[11]。因此,本研究试图寻找新型药物用于治疗白血病,深入分析其是否可能通过调控miRNA及下游靶基因表达而发挥作用,旨在为靶向治疗白血病药物的研发提供实验依据。 奇果菌素是从奇果菌中分离的一种多糖,其具有抗炎、抗肿瘤等作用,研究表明奇果菌素可抑制宫颈癌细胞增殖及促进细胞凋亡[12]。研究表明奇果菌素可通过抑制过氧化物酶体增殖物激活受体γ共激活因子1α(PGC1α)与Fra-1/LSF-MMP2/CD44轴之间的相互作用从而抑制肿瘤细胞生长及迁移,还可通过抑制ERK1/2表达从而抑制肿瘤细胞转移[13,14]。本研究结果显示不同浓度的奇果菌素作用于白血病细胞后,细胞抑制率与细胞凋亡率显著升高,提示奇果菌素可抑制白血病细胞增殖及促进其凋亡。研究报道指出细胞分裂是细胞增殖的基础,细胞周期异常与多种恶性肿瘤发生发展有关,p21是细胞周期素依赖性激酶的抑制因子并可负向调控细胞周期进程从而抑制肿瘤细胞生长,CyclinD1可正向调控细胞周期进程从而促进肿瘤细胞生长[15]。本研究结果发现,奇果菌素作用于白血病细胞后可显著促进p21表达而抑制CyclinD1的表达,且呈一定剂量依赖性,随着药物浓度的增加,细胞增殖的抑制作用显著增强,提示奇果菌素可能通过上调p21的表达及下调CyclinD1的表达从而抑制白血病细胞增殖。细胞凋亡在肿瘤发生发展过程中发挥重要作用,目前大多数抗白血病药物的主要作用机制是诱导白血病细胞凋亡,Bcl-2是抑制细胞凋亡的基因,其可通过参与钙离子依赖的蛋白磷酸化过程而发挥作用,Bax是促细胞凋亡的基因,其可通过激活线粒体途径从而发挥作用[16]。本研究结果显示奇果菌素可诱导白血病细胞凋亡,Bax蛋白的表达水平随着奇果菌素浓度的增加而显著升高,而Bcl-2蛋白的表达水平显著降低,提示奇果菌素可能通过上调Bax蛋白表达及下调Bcl-2蛋白表达从而诱导白血病细胞凋亡。 miR-646过表达可通过抑制p-EGFR、p-Akt蛋白表达从而降低肺癌细胞增殖能力[17]。研究表明miR-646过表达可通过抑制表皮生长因子受体(EGFR)通路激活从而降低骨肉瘤细胞增殖及迁移能力[18]。本研究结果显示奇果菌素可显著上调白血病细胞中miR-646的表达,进一步研究显示miR-646过表达可显著降低白血病细胞增殖能力,同时可促进细胞凋亡,抑制CyclinD1、Bcl-2蛋白表达,促进p21、Bax蛋白表达,提示奇果菌素可能通过上调miR-646表达而抑制白血病细胞增殖及诱导细胞凋亡,其可通过上调p21、Bax的表达及CyclinD1、Bcl-2的表达而发挥作用。本研究发现奇果菌素可抑制ADAM17表达,研究表明沉默ADAM17表达可抑制乳腺癌细胞增殖,还可降低胰腺癌细胞迁移能力[19,20]。本研究通过TargetScan预测显示ADAM17可能是miR-646的靶基因,本研究通过双荧光素酶报告实验证实miR-646可靶向结合ADAM17,并可负性调控ADAM17的表达及活性,进一步研究显示干扰ADAM17表达对白血病细胞增殖具有明显抑制作用,并可促进细胞凋亡,提示奇果菌素可能通过上调miR-646表达而抑制ADAM17表达从而调控白血病细胞增殖及凋亡。为验证上述推测,本研究在白血病细胞中转染miR-646抑制剂后用奇果菌素处理,结果发现细胞抑制率与细胞凋亡率均明显降低,促进CyclinD1、Bcl-2蛋白表达,而抑制p21、Bax蛋白表达,提示抑制miR-646表达可逆转奇果菌素对白血病细胞增殖、凋亡的作用。研究结果证实奇果菌素可通过上调miR-646表达而抑制ADAM17表达从而抑制白血病细胞增殖,诱导细胞凋亡。 综上所述,奇果菌素可通过上调miR-646表达及下调ADAM17表达从而发挥抗白血病的作用,为奇果菌素在白血病的治疗方面的应用提供理论依据。但奇果菌素是否可通过调控其他基因表达或相关信号通路尚需进一步研究。
