基于mtDNA COI基因序列的中国柑橘果实蝇的分子鉴定及柑橘大实蝇种群的遗传多样性分析
2020-03-03崔中翌刘一鹏司品法
崔中翌, 周 琼, 刘一鹏, 司品法, 汪 洋
(湖南师范大学生命科学学院, 长沙 410081)
果实蝇是双翅目(Diptera)实蝇科(Tephritidae)昆虫的统称,是双翅目最大的类群(Aluja, 1999),主要危害水果和蔬菜,对我国的果蔬生产及其出口贸易产生严重影响。世界已知实蝇大约有500个属4 500多种(吴佳教等, 2009),主要分布在亚洲的热带、亚热带和暖温带地区(梁亮等, 2011)。目前已报道的危害我国柑橘的实蝇种类主要包括柑橘大实蝇Bactrocera(Tetradacus)minax、桔小实蝇B.dorsalis和蜜柑大实蝇B.tsuneonis(张宏宇等, 2009; 王俊伟等, 2009; 尤克西等, 2012; Zhangetal., 2018),均隶属于离腹寡毛实蝇属Bactrocera。柑橘大实蝇的寄主仅限于柑橘类果实,对我国柑橘产业造成了巨大的经济损失,该虫在中国的分布包括四川、贵州、湖南、湖北、云南、重庆等省市(Zhang, 1989; Liangetal., 1993; 汪兴鉴和罗禄怡, 1995),在国外分布于印度、尼泊尔、不丹等国家(Vergheseetal., 2004; Dorjietal., 2006)。柑橘大实蝇首次在1940年由Chen(1940)在四川记录到,之后1957年扩增到了中国5个省份,1966年扩增到6个省份,1995年继续扩增到8个省,柑橘大实蝇随柑橘的危害扩散迅速,至今其起源中心与路径还未知(Hongetal., 2019)。
线粒体DNA(mtDNA)是一种在物种进化中相对保守的DNA,一般为母系遗传,无内含子和重复序列且进化速率相对较快;同时,插入和缺失等现象也很少发生(Lin and Danforth, 2004)。朱诗禹等(2017)利用MEGA和DnaSP软件对大豆食心虫Leguminivoraglycinivorella线粒体COⅡ基因片段进行了不同地理种群的遗传分化分析。刘慎思等(2014)利用线粒体COI基因上一段648 bp的序列片段对离腹寡毛实蝇属Bactrocera的21种常见实蝇种类进行了识别与鉴定,结果表明实蝇种内的遗传距离平均0.0043,种间遗传距离平均0.1540,种内、种间遗传距离没有重合区域。Ball等(2005)将mtDNA COI上一段630 bp的片段作为靶标对蜉蝣目(Ephemeroptera)的昆虫进行研究,得出种内和种间序列的平均差异分别为1%和18%的结论,证明了基于COI基因的DNA条形码能有效地区分蜉蝣目昆虫。COI基因除了用于鉴定物种和研究系统发育之外,还可以进行遗传多样性分析。施伟和叶辉(2004)利用线粒体COI基因的601 bp片段研究了云南5个地区桔小实蝇种群的遗传学关系,发现这5个地理种群间遗传分化程度比较低,没有显示出明显的地理分布族群。马英等(2018)利用约600 bp的COI基因片段对青海省的44种蚤类进行了鉴定分析,得出蚤种内遗传距离在0.01%~2.90%之间,种间遗传距离在4%~12%之间。王蒙等(2014)利用线粒体COI基因561 bp的片段对31个种群的桔小实蝇进行了种群遗传分化的研究,推测出了新发生地区桔小实蝇的来源。Leblanc等(2015)研究21个Bactrocerakohkongiae(桔小实蝇复合种)的样本, COI基因(1 464 bp)单倍型网络图显示出了高遗传多样性。
本研究采用DNA条形码技术,对采自我国不同省份和地区的柑橘果实蝇种群进行鉴定,并对为害柑橘的实蝇各类群的亲缘关系和系统发育、以及种内和种间竞争关系进行了分析,为监测危害我国柑橘的实蝇种类(以柑橘大实蝇为主)和分布,研究柑橘大实蝇种群的扩散机理及其防控提供依据。
1 材料与方法
1.1 供试虫源
本研究的实蝇样本分别采自根据文献记载的我国有柑橘大实蝇分布的8个省市16个地区的柑橘园(地理种群),共28个种群(表1),寄主植物包括蜜桔Citrusreticulate, 温州蜜柑Citrusunshiucv. Miyagawa, 蜜柚Citrusmaxima, 脐橙Citrussinensiscv. Navel, 纽荷尔脐橙Citrussinensiscv. Newhall, 椪柑Citrusreticulatacv. Ponkan, 酸橙Citrusaurantium, 甜橙Citrussinensis, 大红甜橙Citrussineniscv. Dahong, 天草Amakusa (橘类与橙类多亲本的复合杂交品种citrus hybrid), 冰糖橙Citrussinensiscv. Bingtang, 狮头柑Citrusspeciosacv. Shitougan和丑橘(不知火)Shiranuhi [Citrusreticulata×(C.reticulata×C.sinenesis)]共13种(或品种)。测试样本共535头,采集方法主要为摘取柑橘树上虫果并解剖获取幼虫和卵,部分样本为柑橘园网捕、黄板或引诱剂诱捕的成虫,以及柑橘园土壤中挖取的蛹,具体采样信息如表1所示。 所采集供测试的果实蝇样本置于75%乙醇中,常温保存备用。 所有标本均存放于湖南师范大学生命科学学院。
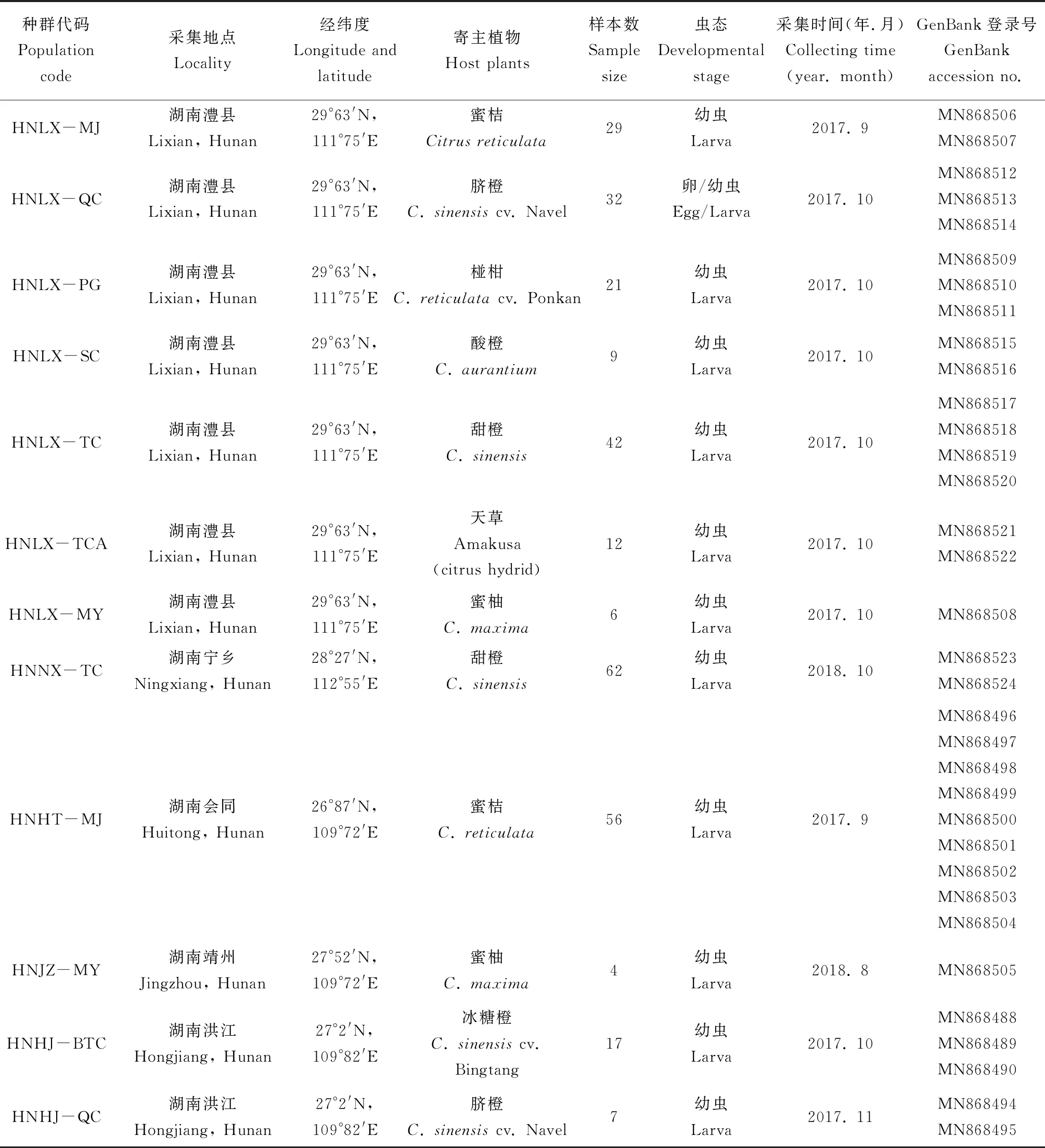
表1 供试柑橘果实蝇样本信息
续表1 Table 1 continued
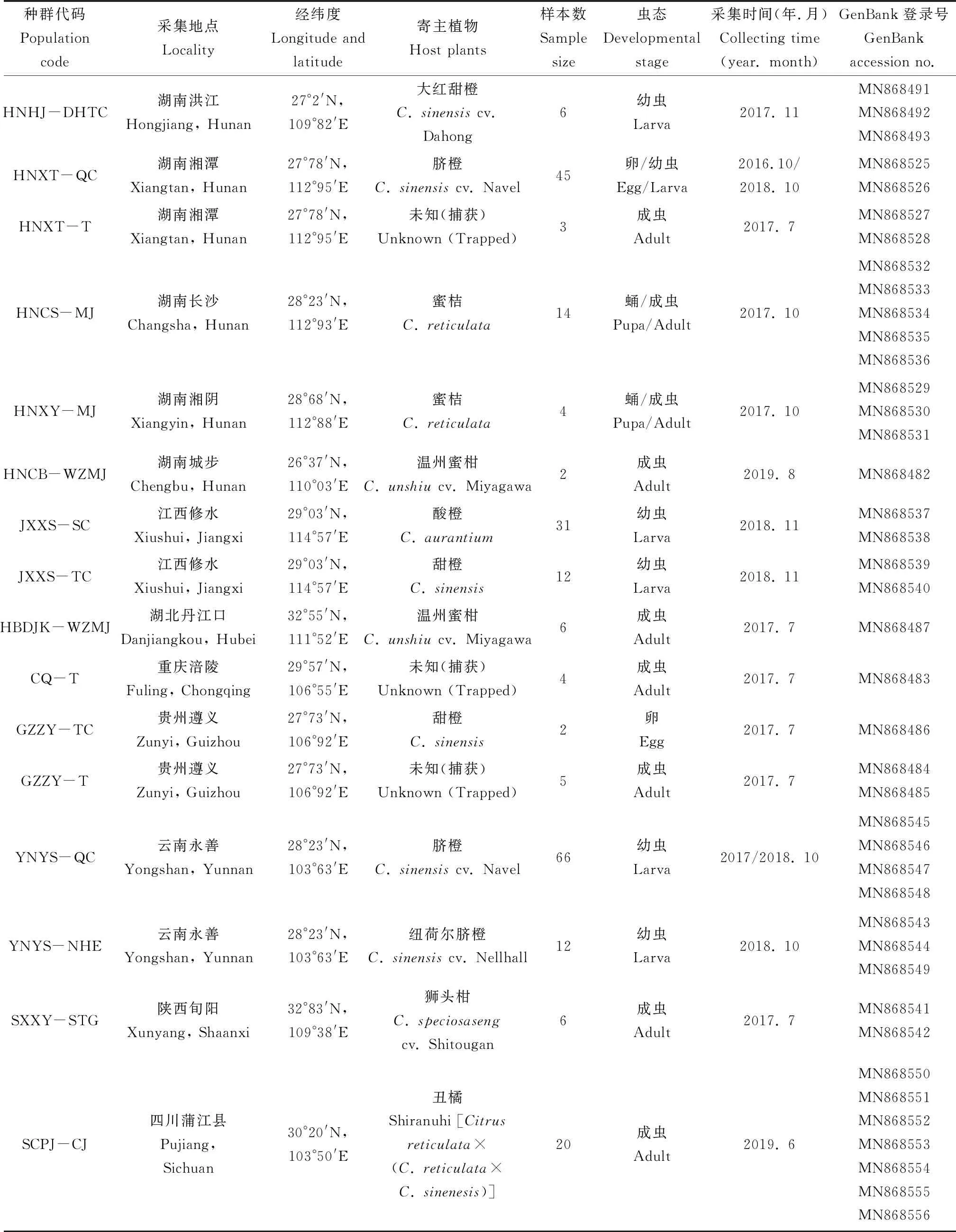
种群代码Populationcode采集地点Locality 经纬度Longitude andlatitude寄主植物Host plants样本数Samplesize虫态Developmental stage采集时间(年.月)Collecting time(year. month)GenBank登录号GenBankaccession no.HNHJ-DHTC湖南洪江Hongjiang, Hunan27°2'N,109°82'E大红甜橙C. sinensis cv. Dahong 6幼虫Larva2017. 11MN868491MN868492MN868493HNXT-QC湖南湘潭Xiangtan, Hunan27°78'N,112°95'E脐橙C. sinensis cv. Navel45卵/幼虫Egg/Larva2016.10/2018. 10MN868525MN868526HNXT-T湖南湘潭Xiangtan, Hunan27°78'N,112°95'E未知(捕获)Unknown (Trapped)3成虫Adult2017. 7MN868527MN868528HNCS-MJ湖南长沙Changsha, Hunan28°23'N,112°93'E蜜桔C. reticulata14蛹/成虫Pupa/Adult2017. 10MN868532MN868533MN868534MN868535MN868536HNXY-MJ湖南湘阴Xiangyin, Hunan28°68'N,112°88'E蜜桔C. reticulata4蛹/成虫Pupa/Adult2017. 10MN868529MN868530MN868531HNCB-WZMJ湖南城步Chengbu, Hunan26°37'N,110°03'E温州蜜柑C. unshiu cv. Miyagawa2成虫Adult2019. 8MN868482JXXS-SC江西修水Xiushui, Jiangxi29°03'N,114°57'E酸橙C. aurantium31幼虫Larva2018. 11MN868537MN868538JXXS-TC江西修水Xiushui, Jiangxi29°03'N,114°57'E甜橙C. sinensis12幼虫Larva2018. 11MN868539MN868540HBDJK-WZMJ湖北丹江口Danjiangkou, Hubei32°55'N,111°52'E温州蜜柑C. unshiu cv. Miyagawa6成虫Adult2017. 7MN868487CQ-T重庆涪陵Fuling, Chongqing29°57'N,106°55'E未知(捕获)Unknown (Trapped)4成虫Adult2017. 7MN868483GZZY-TC贵州遵义Zunyi, Guizhou27°73'N,106°92'E甜橙C. sinensis2卵Egg2017. 7MN868486GZZY-T贵州遵义Zunyi, Guizhou27°73'N,106°92'E未知(捕获)Unknown (Trapped)5成虫Adult2017. 7MN868484MN868485YNYS-QC云南永善Yongshan, Yunnan28°23'N,103°63'E脐橙C. sinensis cv. Navel66幼虫Larva2017/2018. 10MN868545MN868546MN868547MN868548YNYS-NHE云南永善Yongshan, Yunnan28°23'N,103°63'E纽荷尔脐橙C. sinensis cv. Nellhall12幼虫Larva2018. 10MN868543MN868544MN868549SXXY-STG陕西旬阳Xunyang, Shaanxi32°83'N,109°38'E狮头柑C. speciosasengcv. Shitougan6成虫Adult2017. 7MN868541MN868542SCPJ-CJ四川蒲江县Pujiang,Sichuan30°20'N,103°50'E丑橘Shiranuhi [Citrusreticulata×(C. reticulata×C. sinenesis)]20成虫Adult2019. 6MN868550MN868551MN868552MN868553MN868554MN868555MN868556
1.2 mtDNA COI基因信息来源
所使用的COI基因信息来自NCBI数据库以及本研究中分析获得的果实蝇序列(登录号见表1),包括柑橘大实蝇B.minax(GenBank登录号: HM776033.1)(Zhangetal., 2014)、桔小实蝇B.dorsalis(GenBank登录号: DQ845759.1)(徐浪等, 2007)、蜜柑大实蝇B.tsuneonis(GenBank登录号: KM024421.1)(尤欢等, 2014)和瑞丽果实蝇B.ruiliensis(GenBank登录号: KJ753925.1)(尤欢等, 2014)。
1.3 DNA提取
取1.1节所采集的样本于1.5 mL离心管中,其中,1龄幼虫取整虫,2龄幼虫取体前1/2长,3龄幼虫取体前1/3长,蛹取整体,卵取整粒,成虫取一足。分别用直径4.5 mm小钢珠在混合冷冻混合球磨仪(RETSCH MM400)中震碎后,采用天根血液/细胞/组织基因组DNA提取试剂盒(TIANamp Genomic DNA Kit)提取。
1.4 PCR扩增
将1.3节中提取的DNA作为模板,以DNA条形码COI基因的通用型引物(Armstrong and Ball, 2005)进行PCR扩增,LCO1490(5′-GGTCAACAAA TCATAAAGATATTGG-3′)作为上游引物,HCO2198(5′-TAAACTTCAGGGTGACCAAAAAATCA-3′)作为下游引物。反应体系(25 μL):水7.5 μL, 2×Es Taq MasterMix (Dye) 12.5 μL, 上下游引物(10 μmol/L)各1 μL, DNA模板3 μL。扩增程序: 94℃预变性10 min; 94℃ 30 s, 55℃ 30 s, 72℃ 40 s, 32个循环;最后72℃ 10 min。PCR扩增反应在PCR扩增仪(Eppendorf AG 22331 Hamburg)上进行。取3 μL PCR扩增产物经1.0%琼脂糖凝胶电泳检测,最后在Gel-Doc Universal Hood II型凝胶成像系统(Bio-Rad Laboratories)下分析电泳结果。
1.5 PCR产物纯化、测序及序列分析
对1.4节中的PCR扩增产物进行纯化,送深圳华大基因科技有限公司(北京)和铂尚生物技术有限公司(上海)测序。序列采用MEGA 7.0软件进行整合对齐,并利用NCBI数据库对所获得的高质量COI基因序列与已登录的实验昆虫碱基序列(见1.2节)进行同源性对比,DNAstar8.1软件进行序列碱基含量分析。对分别采自云南永善,江西修水以及湖南会同、澧县、宁乡、湘潭和洪江7个地区的43个柑橘虫果内的所有实蝇样本(共343头幼虫或卵)的COI序列,利用MEGA 7.0软件中 Kimura 2-parameter model(Kimura 双参数模型计算)进行种内及种间遗传距离分析。利用DnaSP6.11.01软件分析计算28个种群的果实蝇COI基因单倍型分布、单倍型多样性和核苷酸多样性,并进行Tajima’sD中性检验。使用MEGA 7.0软件中的最大似然(maximum likelihood, ML)法和邻接(neighbor-joining, NJ)法与数据库中现有实蝇种类的COI基因序列一同构建系统进化树,使用Kimura 2-parameter模型建树,Bootstrap方法检验置信度,重复1 000次。使用GraphPad Prism 5.0 软件分析单个柑橘虫果内实蝇数量并作图。用TCS 1.21软件建立单倍型网络树并用PowerPoint 15.37 美化单倍型网络树(图3)。
2 结果
2.1 柑橘果实蝇mtDNA COI的序列特征分析
本研究中获得的所有实蝇的COI基因碱基序列(GenBank登录号见表1)与NCBI数据库中已知的相对应实蝇COI基因片段序列的一致性为99%~100%,且没有碱基插入或缺失现象。使用MEGA7.0软件将所有实蝇COI基因序列进行整合比对,修剪成长度505 bp的片段。使用DNAstar8.1软件对序列进行碱基含量分析,结果表明,所测全部COI基因序列中碱基A, G, T和C的平均含量分别为27.82%, 16.53%, 29.69%和25.95%,A+T的含量为57.51%,C+G含量为42.49%。保守位点C(conserved sites)378个,变异位点V(variable sites)127个,简约信息位点Pi(parsimony information sites)101个,单核苷酸多态性(single nucleotide polymorphisms, SNPs)位点26个。
2.2 柑橘果实蝇种类鉴定
所采集的535头柑橘实蝇中,经COI基因的鉴定,发现其中有520头柑橘大实蝇(占97.20%),10头桔小实蝇(占1.87%),4头蜜柑大实蝇(占0.75%)和1头瑞丽果实蝇(占0.18%)。
柑橘大实蝇的种内遗传距离为0.0000~0.0140,平均为0.0033;桔小实蝇种内遗传距离为0.0000~0.0120,平均为0.0056;蜜柑大实蝇种内遗传距离为0。
4种柑橘果实蝇的种间遗传距离为0.0264~0.2410,平均为0.2094。其中,柑橘大实蝇与蜜柑大实蝇的种间遗传距离较近,为0.0985~0.1127,而与桔小实蝇和瑞丽果实蝇的种间遗传距离分别为0.2212~0.2410和0.2180~0.2290;同时,桔小实蝇与瑞丽果实蝇的遗传距离最近,为0.0264~0.0306,而与蜜柑大实蝇种间遗传距离较远,为0.2093~0.2180。
2.3 柑橘果实蝇的系统发育分析
用最大似然法构建基于COI基因序列的柑橘果实蝇的系统发育树(图1)。可以看出, 4种柑橘果实蝇分属于两大支,其中柑橘大实蝇与蜜柑大实蝇聚为一大支,桔小实蝇与瑞丽果实蝇聚为另一大支。
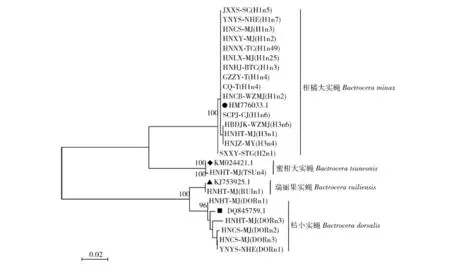
图1 最大似然法构建基于COI序列的柑橘果实蝇的系统发育树
2.4 同一柑橘虫果内的实蝇种类及其遗传距离
对采自云南永善、江西修水、以及湖南会同、澧县、宁乡、湘潭和洪江7个地区的43个柑橘虫果内的343头幼虫或卵进行分析鉴定。发现43个虫果内都有柑橘大实蝇,其中,有39个虫果内的幼虫或卵均为柑橘大实蝇,占所调查虫果的90.7%,其余4个虫果内既有柑橘大实蝇也有其他实蝇(图2),仅占所调查虫果的9.3%(4/43)。单个柑橘虫果内的实蝇数量在4~22头之间。
在39个仅有柑橘大实蝇为害的虫果中,有26个虫果内柑橘大实蝇的个体间遗传距离均为0.0000,占大多数(66.67%);有13个虫果(占比33.33%)的柑橘大实蝇个体间遗传距离范围在0.0000~0.0080。其中遗传距离为0.0020, 0.0040和0.0080的柑橘大实蝇COI基因序列之间分别有1, 2和4个碱基的差异。
其余4个有不同种类的实蝇为害的虫果,均来自湖南会同,虫果内除柑橘大实蝇外,还有桔小实蝇、蜜柑大实蝇或瑞丽果实蝇。这些虫果内的柑橘大实蝇个体间遗传距离为0.0000的有3个(图2: 虫果编号21, 22和23),仅1个虫果(图2: 虫果编号18)的柑橘大实蝇个体间有不同的遗传距离(遗传距离分别为0.0020, 0.0080和0.0100)。
2.5 柑橘大实蝇不同地理种群的遗传多样性
将经过分子鉴定,已明确为柑橘大实蝇的505 bp的COI基因序列,用DnaSP分析各种群的单倍型个数,结果发现,16个地区28个种群的520头柑橘大实蝇,可以分成17个单倍型(表2)。
COI基因序列的单倍型多样性和核苷酸多样性分析显示,17个单倍型中,有8个共享单倍型(H1, H2, H3, H4, H5, H6, H7, H8),9个单倍型为各种群独有(H9, H10, H11, H12, H13, H14, H15, H16, H17),其中四川蒲江寄主为丑橘的柑橘大实蝇种群中单倍型数量最多(7个),其次是湖南会同寄主为蜜桔的种群,单倍型有5个,湖南澧县(寄主为椪柑、脐橙、甜橙)、云南永善脐橙、湖南洪江(寄主为冰糖橙、大红甜橙)、湖南长沙蜜桔、湖南湘阴蜜桔种群中的单倍型种类数量都在3种及以上。柑橘大实蝇28个种群间的单倍型多样性、核苷酸多样性以及核苷酸平均差异数等遗传多样性评价参数的变动范围较大:单倍型多样性范围为0.237~0.833,其中湖南湘阴寄主为蜜桔的柑橘大实蝇单倍型多样性最高;核苷酸多样性范围为0.00049~0.00543,其中湖南会同寄主为蜜桔的柑橘大实蝇核苷酸多样性最高;核苷酸平均差异数变动范围为-1.28172~2.74006,其中湖南会同蜜桔种群核苷酸多样性以及核苷酸平均差异最高。Tajima’sD中性检验结果显示,大部分为正值,湖南会同蜜桔种群的Tajima’sD为2.74006,达到极显著性检验标准(P<0.01),其他种群的Tajima’sD均无显著性,表明柑橘大实蝇种群总体上达到一个动态平衡,未经历明显的种群扩张。

图2 单个柑橘虫果内的实蝇种类、数量及遗传距离
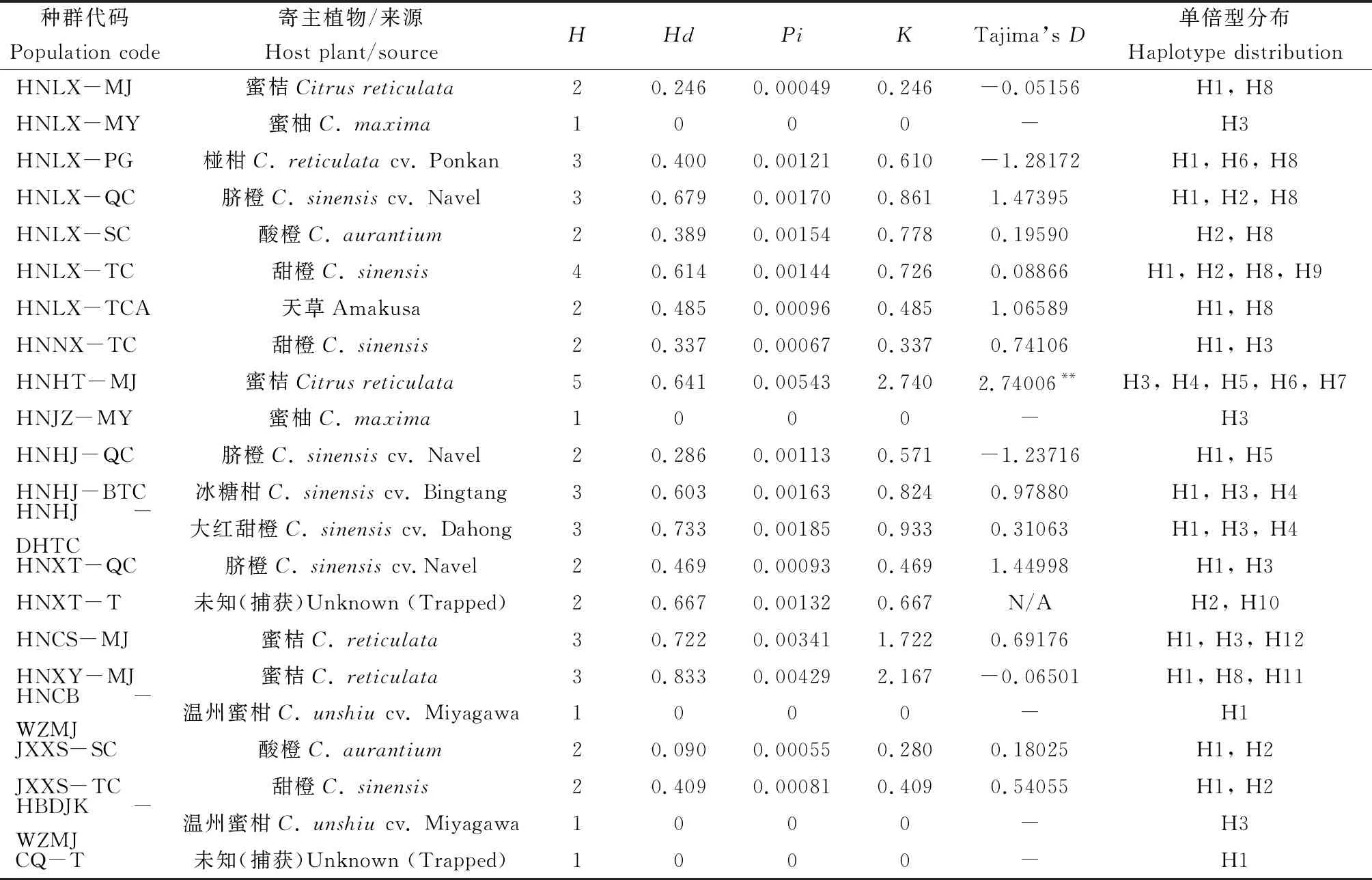
表2 柑橘大实蝇不同地理种群COI基因单倍型多样性、核苷酸多样性分析及中性检验
续表2 Table 2 continued
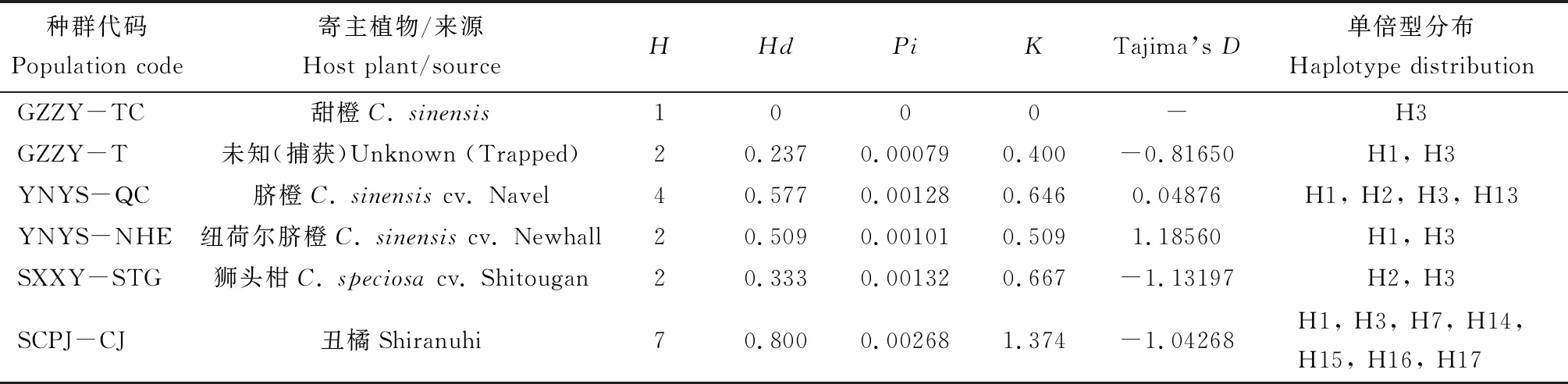
种群代码Population code寄主植物/来源Host plant/sourceHHdPiKTajima’s D单倍型分布Haplotype distributionGZZY-TC甜橙C. sinensis1000-H3GZZY-T未知(捕获)Unknown (Trapped)20.2370.000790.400-0.81650H1, H3YNYS-QC脐橙C. sinensis cv. Navel40.5770.001280.6460.04876H1, H2, H3, H13YNYS-NHE纽荷尔脐橙C. sinensis cv. Newhall20.5090.001010.5091.18560H1, H3SXXY-STG狮头柑C. speciosa cv. Shitougan20.3330.001320.667-1.13197H2, H3SCPJ-CJ丑橘Shiranuhi70.8000.002681.374-1.04268H1, H3, H7, H14,H15, H16, H17
种群代码见表1。For population codes, see Table 1.H: 单倍型数量Number of haplotypes;Hd: 单倍型多样性Haplotype diversity;Pi: 核苷酸多样性Nucleotide diversity;K: 核苷酸平均差异数Average number of nucleotide difference; N/A: 单一样本,不适宜计算遗传多样性One sample only, the genetic diversity is not applicable.**P<0.01.
2.6 柑橘大实蝇不同地理种群单倍型网络
对获得的COI基因序列采用TCS软件进行分析,结果发现,所采获的柑橘大实蝇共有17个单倍型,网络图呈单一散射分布(图3)。其中,单倍型H1位于网络图中心,其他单倍型围绕分布,大部分低频单倍型仅由一到两步突变而成。17个单倍型中,单倍型H1出现频率最高(229/520),占所有检测个体的44.04%,其次是单倍型H3(94/520),占所有检测个体的18.08%, H1和H2均出现在所检测的15个地区中的11个地区种群中。单倍型H11和H12与其他单倍型相对遗传距离较远,具有明显的分支。各单倍型散乱分布,缺乏明显地理分布格局,且有很多共享单倍型。
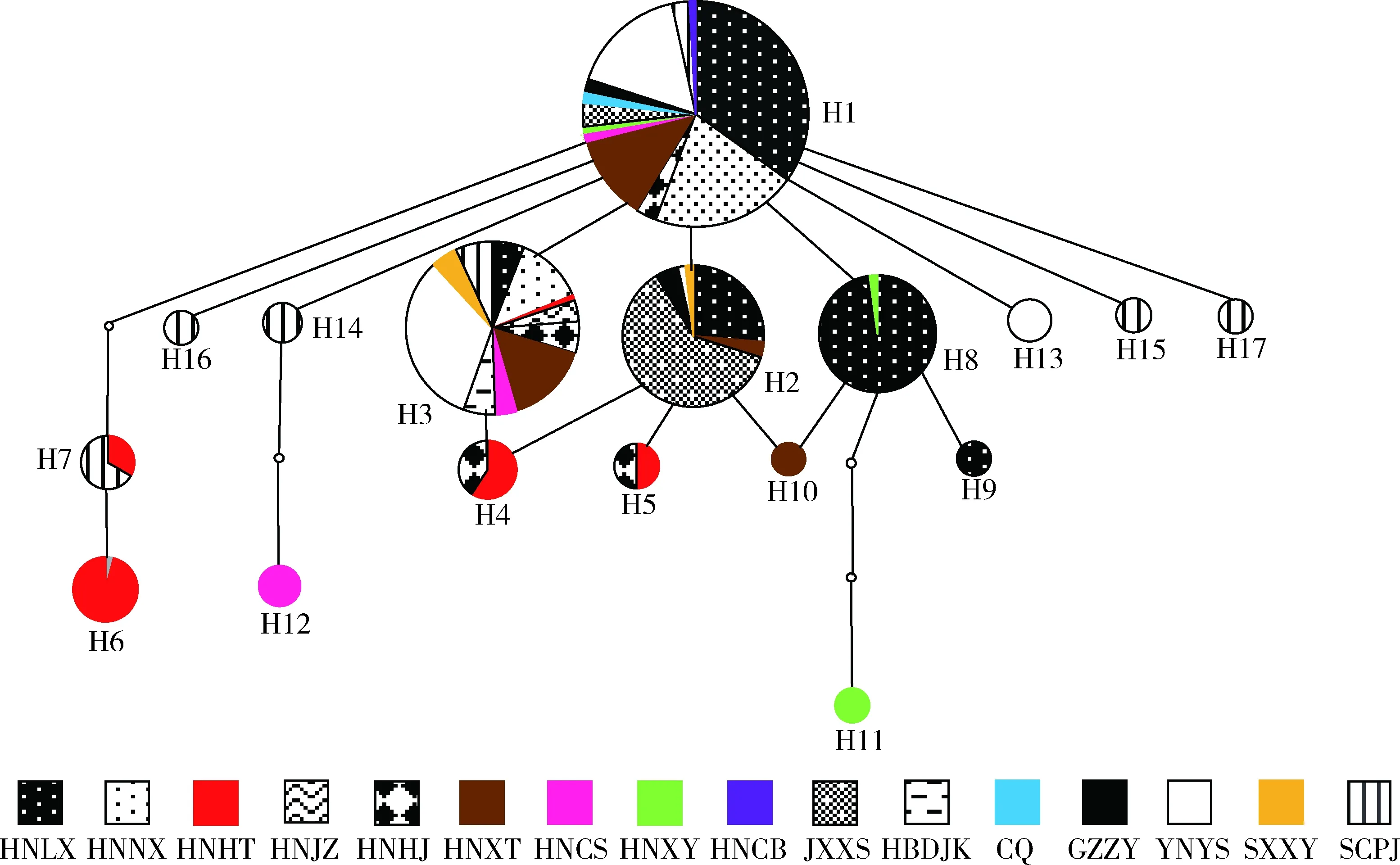
图3 基于COI基因序列的不同地区柑橘大实蝇单倍型网络
2.7 柑橘大实蝇不同地理种群聚类分析
邻接法(NJ)建立的柑橘大实蝇地理种群系统发育树拓扑结构(图4)与最大似然法(ML)的(图1)基本一致,表明拥有同种共享单倍型柑橘大实蝇的地理种群基本汇在一起。不同种群单倍型在系统发育树上有一定的地理分布格局,但并非所有地理种群都聚在一起。
分析还发现,不同地区相同寄主的柑橘大实蝇种群在聚类分析中没有明显关联,而且,同一地区的不同寄主之间以及与相同寄主之间在聚类分析中也未呈现明显规律。说明柑橘大实蝇COI基因的遗传多样性与其对寄主植物的选择没有明显相关。

图4 邻接法构建基于mtDNA COI基因序列的28个柑橘大实蝇地理种群单倍型系统进化树
3 结论与讨论
本研究以柑橘实蝇的卵、幼虫(1-3龄)、蛹以及成虫的足为模板,对我国8省份16个地区危害柑橘的实蝇种类进行了COI基因的分子鉴定分析,发现所调查地区危害柑橘的实蝇主要为柑橘大实蝇,另外还有桔小实蝇、蜜柑大实蝇和瑞丽果实蝇。4种实蝇COI基因所测片段碱基组成与Muraji和Nakahara(2001)研究的实蝇mtDNA碱基组成具有(A+T)偏好性一致。本研究结果符合Hebert等(2003)提出的DNA条形码种间遗传距离在0.02之内,种间差异是种内差异10倍的有效性检测标准;遗传距离分析结果也符合Meier等(2008)提出的双翅目种内遗传距离平均1.3%,种间遗传距离平均10.1%。
瑞丽果实蝇为国内首次在柑橘中发现。在调查解剖统计蜜桔内的柑橘大实蝇时,我们发现1头较小的善弹跳的实蝇幼虫(柑橘大实蝇幼虫没有弹跳习性),通过COI基因片段与NCBI数据库中的序列对比(BLAST),与瑞丽果实蝇(GenBank登录号: KJ753925.1)有很高的核苷酸序列一致性(99%),遗传距离为0.0040,而与Bactrocerathailandica(GenBank登录号: KF659779.1)的遗传距离为0.0100,以遗传距离最近的作为鉴定物种,故确定为瑞丽果实蝇。该种为肖枢等(2008)报道在中缅边界发现,原分布于印度、泰国、老挝、斯里兰卡和美国加州等地;Wang等(2008)在云南瑞丽采集并鉴定了种类。瑞丽果实蝇寄主不详(Wangetal., 2008; 肖枢等, 2008; 梁亮等, 2011; 许桓瑜等, 2015),其与泰国的B.thailandica具有很高的相似性,两者在形态上的主要区别是缝后黄侧条的长度和胫节颜色稍有不同(Wangetal., 2008; 梁亮等, 2011)。由于DNA条形码目前对区分实蝇复合种并不理想(刘慎思等, 2012),而且本研究该种仅采到1头幼虫,故尚不能对其是同种异名还是复合种下结论。
从理论上来说,高水平的遗传多样性有助于外来物种定殖与扩散(施雯等, 2010)。高立志等(2016)利用微卫星引物对采自7省市的18个柑橘大实蝇种群进行了标记、遗传地理距离分析和Mantal检测,发现18个地理种群间的遗传距离较大(0.004~0.264),存在着较大的遗传分化。Hong等(2019)利用线粒体基因nad4片段同样也研究了柑橘大实蝇的遗传多样性以及种群结构,揭示出柑橘大实蝇具有较高的遗传多样性。本研究结果支持以上结论,发现8个省市的28个柑橘大实蝇地理种群具有17个单倍型,在系统聚类树中的分布具一定的地区聚集的分布格局,表明各地理种群间的遗传分化程度较高,但也有少量种群并未与同一地理种群聚类,追究其原因,可能与未经检疫的带虫柑橘在不同地区间的运输和销售引起的被动扩散有关。同时,不同地理种群的相同寄主柑橘品种内的柑橘大实蝇在系统聚类树中并没有聚在一起,同一地理种群的相同寄主和不同寄主的柑橘大实蝇也未显示明显聚类的规律,提示柑橘大实蝇的种内遗传分化与地理分布有关,而与对寄主的选择没有明显的关联。柑橘大实蝇地理种群的高水平的遗传多样性,提示其有较强的扩大危害范围的风险。
竞争是指多个个体为了同一资源而产生的争夺(李国清和慕莉莉, 2006)。引起种间竞争的主要因素是生态位的重叠(马骏等, 2009)。广食性的桔小实蝇与番石榴实蝇(刘晓飞等, 2014)、瓜实蝇与南亚果实蝇(张金龙等, 2017)之间存在种间竞争关系。柑橘大实蝇为寡食性,仅危害芸香科柑橘属植物果实,其一年发生1代(汪兴鉴和罗禄怡, 1995);桔小实蝇是广食性,寄主多样包括芒果、柑橘、杨桃等,其一年3~5个世代,在不同地区完成世代有所不同(王小蕾和张润杰, 2009);蜜柑大实蝇与柑橘大实蝇有着相近的外型、生活史、行为与寄主(Zhangetal., 2018),其二者外型区别主要在于有无前翅上鬃,有无肩板鬃中对(汪兴鉴和罗禄怡, 1995);瑞丽果实蝇的生活史周期不详,Wang等(2008)在2003-2006年的5-10月期间诱捕得到其标本并鉴定。这4种实蝇均为害柑橘果实并具有明显的生态位重叠,但仅柑橘大实蝇在我们所调查地区是危害柑橘的绝对优势实蝇种类,表明这4种实蝇的生存竞争中,柑橘大实蝇在我们所调查地区具有强大的竞争优势,其机制尚待研究。
一些实蝇在产卵的同时,雌成虫会留下寄主标记信息素,寄主标记信息素主要作用通常就是防止其他雌性在此处再次产卵,阻止过度寄生(李国清和慕莉莉, 2006; Chesetoetal., 2017)。Hågvar(1988)对寄生于桃蚜上的两种蚜茧蜂进行了研究,认为寄主标记信息素对昆虫种间竞争的影响常见于近缘种不同个体之间。实蝇科的多种昆虫具有寄主标记行为,如桔小实蝇(赵静等, 2014)、地中海实蝇Ceratitiscapitata(Arredondo and Díazfleischer, 2006)和墨西哥实蝇Anastrephaludens(Alujaetal., 2009)等。本研究结果发现,采自不同地区的43个虫果中,有66.67%(26/43)的同一虫果内柑橘大实蝇遗传距离均为0.0000;有30.23%(13/43)虫果内有大小差异的不同龄期的柑橘大实蝇幼虫且遗传距离大于0.0000,说明可能存在部分多头柑橘大实蝇雌虫先后在同一果实中产卵的情况;同时,仅有9.3%(4/43)的虫果内同时有不同种类实蝇(柑橘大实蝇、蜜柑大实蝇、桔小实蝇或瑞丽果实蝇)寄生。上述结果提示,柑橘大实蝇可能也存在寄主标记行为,但其产卵标记信息物的作用效果有一定限度。
我们在调查中还发现,在柑橘果园中利用食诱剂、诱球和黄板还可以诱捕到少量的南瓜实蝇Bactroceratau和具条实蝇B.scutellata成虫,但目前尚未在柑橘虫果内获得该两种实蝇幼虫的分子证据,因此,本研究尚不能明确这两种实蝇是否危害健康柑橘果实,推测诱捕到的南瓜实蝇和具条实蝇成虫可能源自橘园周边的瓜果地或落果。
致谢承蒙西南大学王进军教授、华中农业大学张宏宇教授和牛长缨教授、云南农业大学肖春教授和董文霞教授、长江大学桂连友教授、江西修水余春华高级农艺师、云南永善县柯贤江农艺师以及四川成都浦江县周伦高级农艺师提供柑橘大实蝇标本或虫果,贵州遵义市殷元进高级农艺师和湖南省植保站张佳峰农艺师协助采集标本,在此一并表示衷心的感谢!
