食盐添加量对哈尔滨风干肠脂质和蛋白氧化及挥发性化合物形成的影响
2020-02-10温荣欣扈莹莹殷小钰孔保华
温荣欣,扈莹莹,殷小钰,王 妍,孔保华,陈 倩,
(1.东北农业大学食品学院,黑龙江 哈尔滨 150030;2.岛津企业管理(中国)有限公司,辽宁 沈阳 110000)
哈尔滨风干肠是我国北方传统自然发酵肉制品,因其独特的口感与风味深受消费者喜爱。其加工过程需要历经较长时间的风干和发酵,为了抑制其中致病菌和腐败菌的生长和繁殖,保证产品安全性和货架期,需要较高的食盐(主要成分为NaCl)添加量。然而,哈尔滨风干肠发酵周期较长(12~15 d),随着风干和发酵的进行,风干肠中的水分含量不断减少,导致终产品中NaCl含量较高(3.3%~4.0%)[1]。因此,降低干香肠中的NaCl含量受到了越来越多的关注。如若长期摄入高食盐含量的肉制品,会增加患高血压、中风和心血管等疾病的风险[2]。因此,世界卫生组织建议成人每日食盐摄入量低于5 g(<2 g Na/d)[3],《中国居民膳食指南》也推荐成人每日食盐摄入量不超过6 g[4]。然而,现在中国居民平均食盐摄入量达到10.5 g/d。为减少国民食盐摄入量,在2018年4月18日,中国疾病预防控制中心营养与健康所和中国营养学会在北京联合召开了中国食品工业减盐指南研讨会,提出到2030年全国人均每日食盐摄入量降低20%的目标[5]。
食盐是肉制品加工中不可或缺的腌制材料,不仅可赋予产品咸度,增加鲜度,而且对加工特性以及品质特性的形成具有重要的贡献作用[6]。因此,降低食盐添加量的同时保证肉制品的品质及安全性是肉品行业亟待解决的热点问题。关于减盐处理通常采用NaCl替代物和直接减盐的方式[7]。目前,研究多集中在食盐替代物或减盐处理对发酵香肠理化及品质特性的影响[8-9],但其对脂质和蛋白质氧化的影响却鲜有研究,特别是对香肠中挥发性化合物的产生水平和最终产品中香气构成的影响。基于此,本研究采用直接减少哈尔滨风干肠中食盐添加量的方法,考察其对风干肠发酵过程中脂质氧化、蛋白氧化及挥发性化合物生成的影响,为开发低盐哈尔滨风干肠提供理论依据。
1 材料与方法
1.1 材料与试剂
猪后臀肉及猪背脂、食盐、亚硝酸钠、绵白糖、味素、玉泉大曲、香辛料(世一堂干肠料,包含橘皮、砂仁、丁香、肉豆蔻、白芷、桂皮、小茴香)均为食品级 市购;牛血清白蛋白(bovine serum albumin,BSA)、2,4-二硝基苯肼(2,4-dinitrofenylhydrazin,DNPH)、5,5’-二硫代双(2-硝基苯甲酸)(5,5’-dithiobis (2-nitrobenzoic acid),DTNB)、1-苯氨基-8-萘磺酸盐(1-anilinonaphthalene-8-sulfonic acid,ANS)等均为 分析纯。
1.2 仪器与设备
HWS-70BX恒温恒湿箱 天津市泰斯特仪器有限公司;GC-3L小型灌肠机 瑞安市鸿飞机械有限公司;GL-21M高速冷冻离心机 湖南湘仪实验仪器开发有限公司;T6新世纪紫外-可见分光光度计 北京普析通用仪器有限责任公司;F-4500荧光分光光度计 日本日立公司;GCMS-QP2020单四极杆型气相色谱-质谱联用仪 日本岛津公司。
1.3 方法
1.3.1 哈尔滨风干肠的制备
参照Chen Qian等[10]的方法并作适当修改。本实验共制备4 组风干肠,食盐添加量分别为2.5%(对照)、2.0%、1.5%和1.0%。原料肉为猪后臀肉和猪背脂,将原料肉修整后分别用绞肉机在1.5 cm的孔板上绞碎,两者以9∶1的比例混合均匀。按照配方加入1%绵白糖、0.3%味素、0.01%亚硝酸钠、5%水、1%玉泉大曲、0.8%香辛料,并按照实验设计加入相应的食盐。充分搅拌后灌入猪小肠衣中,每根风干肠长度约20 cm,直径约1.5 cm。然后将风干肠悬挂在温度为(25f 2)℃,相对湿度为30%~50%的环境中风干1 d,然后转移到温度为(25f 2)℃,相对湿度为75%~80%的环境中发酵11 d。分别在第0、3、6、9、12天进行取样,测定其脂质和蛋白氧化的程度,并测定第0、6、12天的挥发性化合物的含量。
1.3.2 脂质过氧化值(peroxide value,POV)测定
参照Vareltzis等[11]的方法并略作修改。取切碎的风干肠(2.0 g)与15 mL氯仿-甲醇(2∶1,V/V)溶液混合,高速均质30 s,加入3.0 mL 0.5% NaCl溶液,4 ℃、3 000h g离心10 min。收集下层液相(5.0 mL)与5.0 mL氯仿-甲醇(2∶1,V/V)溶液混合,并加入25.0 µL 30%硫氰酸铵溶液和25.0 µL氯化亚铁溶液(0.4 g氯化钡和0.5 g硫酸亚铁各自溶于50.0 mL水中,混合并2 000h g离心5 min)。室温反应5 min,于500 nm波长处测定吸光度。以还原铁粉作标准曲线,最终POV表示为mmol/kg。
1.3.3 肌原纤维蛋白和肌浆蛋白的提取
分别按照Liu Gang[12]和Chen Qian[13]等的方法在4 ℃条件下提取肌原纤维蛋白和肌浆蛋白。将提取出的2 种蛋白置于4 ℃环境中,在24 h内完成羰基含量、总巯基含量和表面疏水性的测定。以BSA为标准测定2 种蛋白的浓度。
1.3.4 羰基含量的测定
按照Chen Qian等[13]的方法测定蛋白质的羰基含量。取1 mL质量浓度为2 mg/mL的蛋白(肌原纤维蛋白或肌浆蛋白)溶液与1 mL浓度为10 mmol/L的DNPH在室温下反应1 h(每15 min旋涡振荡1 次),对照组以1 mL 2 mol/L HCl溶液取代DNPH。此后,加入1 mL 20%三氯乙酸溶液,8 500h g离心5 min,弃清液,取沉淀。将沉淀物用1 mL乙酸乙酯(1∶1,V/V)溶液洗涤3 次,在37 ℃溶于3 mL 6 mol/L盐酸胍溶液中保温15 min溶解沉淀,然后 8 500h g离心3 min除去不溶物质,在波长370 nm处测定上清液吸光度。使用分子吸光系数22 000 L/(mol•cm),以蛋白质量计算羰基含量,以nmol/mg表示。
1.3.5 总巯基含量的测定
按照Ellman[14]的方法,用DTNB进行测定。取1 mL质量浓度为2 mg/mL的蛋白(肌原纤维蛋白或肌浆蛋白)溶液与8 mL Tris-甘氨酸溶液(pH 8.0,10.4 g/L Tris,6.9 g/L甘氨酸,1.2 g/L EDTA,480.48 g/L尿素)混合并均质。取4.5 mL混合液与0.5 mL Ellman试剂(10 mmol/L DTNB)反应30 min,然后10 000h g离心15 min,取上清液于波长412 nm处测定吸光度。使用分子吸光系数13 600 L/(mol•cm),以蛋白质量计算总巯基含量,并表示为nmol/mg。
1.3.6 蛋白质表面疏水性的测定
按照Chelh等[15]的方法测定肌原纤维蛋白的表面疏水性。取1 mL质量浓度为1 mg/mL的蛋白溶液加入200 µL溴酚蓝(1 mg/mL)溶液混匀,对照组为1 mL pH 7.0的磷酸盐缓冲液(10 mmol/L)加入200 µL溴酚蓝。室温下搅拌10 min,然后3 500h g离心15 min,上清液(稀释10 倍后)在波长595 nm处测定吸光度。表面疏水性以溴酚蓝质量表示,按式(1)计算:
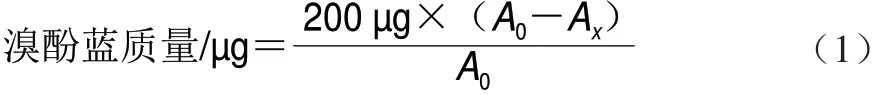
式中:A0为对照组的吸光度;Ax为样品的吸光度。
参照ANS荧光探针法[16]并略作调整,测定肌浆蛋白表面疏水性。取4 mL质量浓度分别为0.02、0.04、0.06、0.08 mg/mL和0.1 mg/mL的肌浆蛋白溶液,分别加入20 µL浓度为8 mmol/L的ANS溶液,充分振荡后在室温下反应20 min,测定荧光强度。荧光测定条件为:激发波长365 nm,发射波长484 nm,狭缝宽5 nm。以荧光强度对蛋白质量浓度作图,斜率即为肌浆蛋白的表面疏水性指数。
1.3.7 挥发性化合物的测定
1.3.7.1 挥发性化合物定性及定量
参照Benito等[17]的方法并略作修改。分别在发酵第0、6、12天取样,采用固相微萃取方法提取挥发性化合物。将切碎的风干肠(3.0 g)放入20 mL顶空样品瓶中,加入4 µL邻二氯苯作为内标物。在45 ℃条件下用50/30 µm DVB/CAR/PDMS萃取头顶空吸附30 min,然后立即将萃取头插入气相色谱-质谱进样口,解吸3 min。
气相色谱条件:InertCapWax毛细管柱(60 mh 0.25 mm,0.25 µm);进样口温度230 ℃;不分流模式进样;以氦气为载体,流速1 mL/min;程序升温:柱初温40 ℃,保持3 min,以5 ℃/min升至200 ℃,再以 10 ℃/min升至230 ℃,保持2 min。
质谱条件:电子电离源;离子源温度230 ℃;质量扫描范围m/z 45~500。
挥发性化合物的鉴定通过比较NIST 14质谱库中的实验质谱,取相似度大于90%作为鉴定结果,并且通过计算标准烷烃(C6~C20)的保留指数,与文献报道结果相比较进行辅助鉴定。挥发性化合物的定量采用内标法,以溶于正己烷的邻二氯苯溶液作为内标物质,定量结果以µg/kg表示。
1.3.7.2 香气活性值(odor activity value,OAV)
参考冯云子[18]的方法,按式(2)计算OAV:

式中:C为挥发性化合物含量/(µg/kg);T为挥发性化合物的气味阈值/(µg/kg)。
1.4 数据统计
实验重复3 次,每组3 个平行,结果表示为f s。数据统计分析采用Statistix 8.1软件包中Linear Models程序进行,差异显著性(P<0.05)分析使用Tukey HSD程序,采用Sigmaplot 12.5软件和Heml 1.0.3.7软件绘图。
2 结果与分析
2.1 食盐添加量对风干肠POV的影响
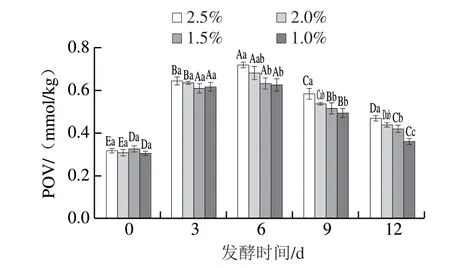
图 1 食盐添加量对风干肠发酵过程中POV的影响Fig. 1 Effect of salt addition on the POV of dry sausages during fermentation
如图1所示,POV在发酵前期(0~6 d)显著增加(P<0.05),说明风干肠中的脂质随发酵时间延长逐渐发生氧化。在发酵后期(6~12 d),POV显著降低 (P<0.05),说明脂质初级氧化产物发生分解,生成次级氧化产物丙二醛以及其他醛、酮、酸等物质。在发酵过程中,风干肠的脂质氧化程度随食盐添加量的增加逐渐增高,其中添加2.5%食盐的风干肠POV最大,这主要是由于NaCl促进了脂质氧化。一方面,NaCl破坏细胞膜结构完整性,促进氧化因子与不饱和脂肪酸的相互 作用[19];另一方面,NaCl促进血红蛋白和肌红蛋白释放铁离子,通过铁离子的催化作用进而促进脂质氧化[20];另外,NaCl可抑制抗氧化酶如过氧化氢酶,谷胱甘肽过氧化物酶和超氧化物歧化酶的活性,从而降低肉制品自身的抗氧化能力[21]。脂质氧化是风干肠在发酵过程中形成特征风味的主要途径,脂质氢过氧化物进一步氧化或降解产生的挥发性化合物是形成风味的主要物质,适当提高氢过氧化物的降解速率有利于加快产品风味的形成。然而,脂质过度氧化易于产生酸败味和哈喇味等不愉快的气味,因此,控制脂质氧化程度对于产品的风味形成至关重要。
2.2 食盐添加量对肌肉蛋白羰基含量的影响

图 2 食盐添加量对风干肠发酵过程中肌原纤维蛋白(A)和 肌浆蛋白(B)羰基含量的影响Fig. 2 Effects of salt addition on the carbonyl contents of myofibrillar proteins (A) and sarcoplasmic proteins (B) in dry sausage during fermentation
如图2所示,发酵初期(0 d),肌原纤维蛋白和肌浆蛋白的羰基含量分别为0.6 nmol/mg和0.9 nmol/mg,随发酵时间的延长,2 种蛋白质的羰基含量显著增 加(P<0.05),这与Wu Haizhou等[22]的研究结果一致。从图2可以看出,肌浆蛋白的氧化程度在发酵第0天大于肌原纤维蛋白,可能因为在肌肉组织中,肌浆蛋白比肌原纤维蛋白更容易与氧气和脂质接触。在发酵末期(12 d),随食盐添加量的增加,肌原纤维蛋白和肌浆蛋白的羰基含量均显著增加(P<0.05),这表明在风干肠发酵过程中NaCl作为促氧化剂,促进了蛋白质中羰基化合物的形成。
在肌肉组织中,蛋白质氧化主要由金属离子 (Fe3+、Cu2+)、肌红蛋白或脂质氧化系统引发[23],NaCl不仅可以增强Fe3+的活性,促进自由基的形成[24],而且结合POV结果可知,NaCl可以促进脂质氧化,由脂质氧化形成的自由基和氢过氧化物也可以攻击氨基酸侧链生成羰基[23]。此外,对于肌原纤维蛋白而言,不同处理组之间的显著差异也可能归因于增加NaCl使离子强度增加,改变了肌原纤维蛋白的溶解度,使其结构变的松散,导致自由基和其他促氧化因子的扩散速度加快[25]。羰基的形成通常会影响肉制品的品质,一些必需氨基酸被氧化破坏后导致肉制品营养价值降低,羰基化合物与非氧化赖氨酸的游离氨基反应形成亚胺键,导致聚集体的形成,降低蛋白质消化率[26]。此外,蛋白质羰基化合物易形成特定的Strecker醛,作为挥发性化合物的重要组分,构成肉制品的独特风味[27]。
2.3 食盐添加量对肌肉蛋白总巯基含量的影响

图 3 食盐添加量对风干肠发酵过程中肌原纤维蛋白(A)和 肌浆蛋白(B)总巯基含量的影响Fig. 3 Effect of salt addition on the total sulfhydryl contents of myofibrillar proteins (A) and sarcoplasmic proteins (B) in dry sausages during fermentation
蛋白质巯基损失也是肉及肉制品中蛋白质氧化的标志之一,巯基被氧化后主要生成蛋白质内或蛋白质间的二硫键以及混合二硫化物[28]。如图3所示,随着发酵的进行,各组风干肠的巯基含量显著降低(P<0.05),这主要是因为含硫氨基酸的巯基容易受到自由基的攻击,易发生交联[24]。肌原纤维蛋白中巯基含量的变化比肌浆蛋白更明显,这与Li Binbin等[29]的研究结果一致,主要是因为构成肌原纤维蛋白的肌球蛋白和肌动蛋白中巯基含量较多,易于优先成为被氧化的目标。各组风干肠的总巯基含量在发酵末期差异显著(P<0.05),这可归因于NaCl对自由基形成和促氧化因子扩散的促进作用,该结果与羰基含量的测定结果一致,进一步证明了NaCl对蛋白质氧化的诱导作用。
2.4 食盐添加量对肌肉蛋白表面疏水性的影响

图 4 食盐添加量对风干肠发酵过程中肌原纤维蛋白(A)和 肌浆蛋白(B)表面疏水性的影响Fig. 4 Effect of salt addition on the surface hydrophobicity of myofibrillar proteins (A) and sarcoplasmic proteins (B) in dry sausages during fermentation
在蛋白质的天然结构中,亲水性氨基酸残基暴露于水相,而疏水性基团通常存在于分子内部[30]。如图4 所示,肌原纤维蛋白和肌浆蛋白的表面疏水性在发酵过程中显著增加(P<0.05),主要因为分子内部的疏水性基团暴露到蛋白质表面,这与Chen Qian等[13]的研究结果一致。在发酵末期(12 d),随食盐添加量的增加,肌原纤维蛋白和肌浆蛋白的表面疏水性均显著增加 (P<0.05)。Kobayashi等[31]研究表明,NaCl有助于鱼糜中蛋白质结构的展开,导致疏水性增大。Wang Guan等[32]发现NaCl浓度升高导致肌球蛋白逐渐解离,暴露疏水性氨基酸残基。表面疏水性的增加会导致蛋白质二级结构和三级结构发生变化,有利于蛋白质发生交联并生成非共价聚集体[33]。此外,埋藏在蛋白质内部疏水性氨基酸的暴露可能易于蛋白酶对特定位点的识别,加速蛋白质的降解并产生更多的肽和游离氨基酸,促进风味物质的形成[34]。蛋白质还可以作为风味物质的载体,暴露的疏水区域也可增加风味化合物的结合位点,影响风味的感知度[35]。
2.5 食盐添加量对风干肠挥发性化合物形成的影响
为了研究不同食盐添加量对风干肠风味形成的影响,以及发酵过程中挥发性物质之间的特征差异,通过气相色谱-质谱对发酵第0(发酵初期)、6(发酵中期)、12天(发酵末期)的样品中挥发性化合物的含量进行测定。在所有样品中共检测出62 种挥发性化合物,包括6 种醛、7 种酮、19 种醇、7 种羧酸、13 种酯、10 种烯烃。如表1所示,随着发酵的进行,挥发性化合物的种类和生成量不断增加,在不同处理组之间存在显著差异(P<0.05)。
源自脂质自氧化和蛋白氧化的挥发性化合物一般具有较低的香气阈值并在风味中占主要地位,但产生量过多时易生成不愉快气味[36]。在风干肠中检测出的直链醛,如己醛、辛醛和壬醛主要由不饱和脂肪酸过氧化反应产生[37],而且这些醛类物质经进一步氧化生成了相对应的酸类物质。如表1所示,第0天检测到壬醛和己酸,其他几种醛类在发酵过程中逐渐产生并积累。发酵末期,醛类化合物的生成量随食盐添加量降低显著减少 (P<0.05),这与获得的POV结果一致,可归因于NaCl的促脂质氧化作用。蛋白氧化产生的羰基易与氨基酸通过美拉德反应和Strecker降解反应生成醛类[27],在风干肠中检测到的苯甲醛和苯乙醛正是由苯丙氨酸经过Strecker降解产生[38]。在发酵末期,食盐添加量的减少显著降低了风干肠中这2 种醛类物质的含量,这与羰基结果一致。
微生物代谢产生的挥发性化合物在风干肠中种类最为丰富,由表1可知,碳水化合物发酵产生的酮、醇和酸均与食盐添加量密切相关。在发酵末期,食盐添加量越低的风干肠,3-羟基-2-丁酮、乙醇和2,3-丁二醇的含量越高。尽管醇类物质的生成量很多,但由于它们香气阈值较高,对风干肠的整体风味贡献不大。然而,由碳水化合物发酵产生的乙酸、丁酸等酸类物质与风干肠特有的成熟香气有关[39]。微生物代谢产生的挥发性化合物含量随食盐添加量降低而呈升高的趋势,该结果表明降低食盐添加量可能减少对相关微生物的抑制作用。1-辛烯-3-醇和2-壬酮是由微生物的脂质不完全β-氧化产生的,由于具有低香气阈值,成为风干肠整体风味的主要贡献者[40]。
乙酯类化合物在酯类化合物中最为丰富,如乙酸乙酯、丁酸乙酯和己酸乙酯等,都是源于微生物的酯化作用[41]。酯类化合物的产生与底物(醇类和酸类)水平以及葡萄球菌的酯酶活性有关[42]。酯类带有果香味,并且它们的香气阈值较低,在风干肠的整体风味中起重要作用[43]。 如表1所示,降低食盐添加量显著增加了酯类化合物的水平,表明低盐处理增强了产酯酶微生物的活性,而且这也可以解释在低盐处理中以相同比例发现的高水平的醇类物质和酸类物质。此外,在风干肠中检测到的少量酮、醇、酯类和所有烯烃类化合物,其主要源于香辛料,如小茴香酮、桉叶油醇、芳樟醇、乙酸香叶酯、α-律草烯和α-蒎烯等。在整个发酵过程中,这些化合物的含量逐渐增加,主要由于风干肠在该过程中水分减少浓缩所致。
通常认为OAV大于1的化合物可以作为主要风味物质,而且OAV越大风味贡献能力越强[18]。图5结果表明,大部分醛类、酮类和酯类物质均具有较高的OAV,而所有的酸类物质和醇类(除1-辛烯-3-醇)物质的OAV均小于1,因此在风干肠整体风味中,醛类、酮类和酯类物质占据重要地位。源于脂质氧化反应的挥发性化合物都具有较高的OAV,而且在发酵末期,源于蛋白质Strecker醛反应的苯乙醛的OAV也大于1。可见脂质氧化和蛋白氧化对风干肠的整体风味都有较大的贡献。

表 1 食盐添加量对风干肠发酵过程中挥发性化合物含量的影响Table 1 Volatile compounds identified and quantified by GC-MS in dry sausages with different levels of salt addition on day 0, 6 and 12 of fermentation µg/kg

续表1 µg/kg

图 5 食盐添加量对风干肠发酵过程中挥发性化合物OAV的影响Fig. 5 Effect of salt addition on the OAV of volatile compounds in dry sausages during fermentation
通过以上结果可知,低盐处理一方面通过降低脂质氧化和蛋白氧化从而减少来自这2 个途径的挥发性化合物的生成量;另一方面增加了由微生物代谢途径产生的挥发性化合物的生成量,改变了风干肠的整体风味构成。另外,前期研究结果表明2.0%食盐添加量的风干肠在感官评价中可接受性最高[1],而且风味指标的分值也超过了2.5%添加量的对照组,说明将食盐添加量减少至2.0%改善了风干肠的整体风味构成。
3 结 论
减少食盐添加量可显著降低风干肠中脂质和蛋白的氧化程度,从而降低了源于这2 种氧化途径的挥发性化合物水平,但是增加了源于微生物代谢作用的化合物水平。结果表明,风干肠的感知香气主要是由具有较高OAV的醛类、酮类和酯类物质组成,整体而言,低盐处理降低了终产品中这些醛类物质的水平,增加了酮类、酸类、酯类和醇类物质的水平,使得风干肠的整体风味发生改变。综上,减少风干肠食盐添加量至2.0%,不仅减少了产品中20%的NaCl,而且改善了产品的风味结构,有助于生产低盐健康的哈尔滨风干肠。
