耐低温生物脱氮机制与对策研究进展
2020-01-15陆浩良田晴朱艳彬张健焦彭博林欢
陆浩良,田晴,朱艳彬,张健,焦彭博,林欢
(东华大学环境科学与工程学院,上海201620)
水中的氮(N)、磷(P)等营养元素的累积会引起富营养化,因此脱氮除磷是污水处理的重要任务。目前城市污水处理厂大多采用A2/O、氧化沟(oxidation ditch)或序批式活性污泥法(SBR)等生物处理工艺,虽然上述工艺能够有效去除污水中的氮磷等元素,但如需保持较高的脱氮除磷效率,需要控制曝气量与投加额外碳源[1]。为了降低运行成本,研究者从这两个角度出发,提出了厌氧氨氧化(anammox)、同步硝化反硝化(SND)等工艺。然而以上所提及的生物处理法中功能菌的生存都依赖于适宜的温度,例如硝化菌的最佳生长温度为20~30℃。由于地理位置和季节的变化,难免有低温环境存在,相关研究表明,长期的低温环境非常不利于微生物的生长代谢和水中污染物的去除,低温会导致微生物群落结构发生改变、生长速率下降、生理特性恶化等,使得出水氨氮和总氮不达标。尤其是在我国的北方地区,以东北地区为例,年均气温约为5℃,低水温使得污水处理厂的出水氨氮和总氮很难达到生物处理的要求。针对上述原因,研究低温条件下高效脱氮、掌握低温硝化菌的脱氮性能,有利于提高污水处理厂的出水效果,具有重要的理论与现实意义。
1 低温微生物
1.1 低温微生物的分类
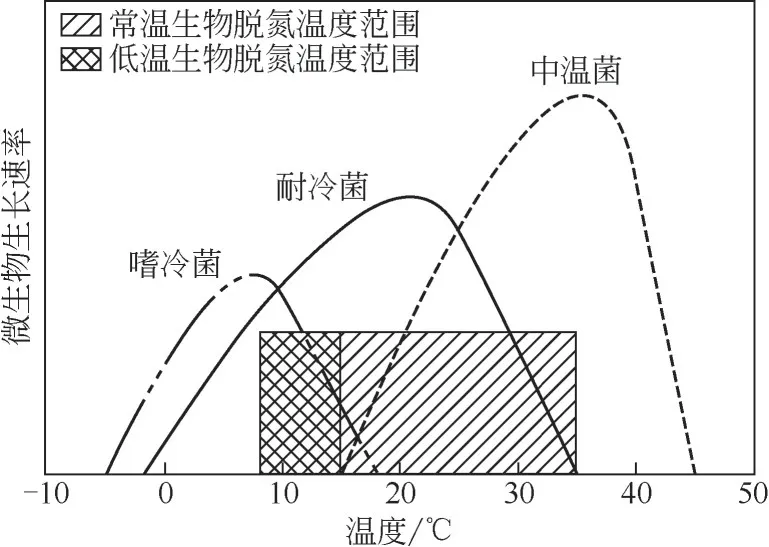
图1 温度对微生物生长的影响
不同种类的微生物适合不同的生长温度,温度对微生物生长的影响如图1 所示[2],通常温度低于15℃便是低温[3]。近年来,各国研究者对地球上各种水生和陆地寒冷环境中的低温微生物进行了多方位的研究,低温微生物的概念也随着研究的深入在不断地完善。自从1887年Forster首次发现在0℃条件下生长的微生物以来,先后出现描述低温微生物的名词多达十几种。目前学术界普遍认可的是Morita[4]于1975 年给出的定义,他将这类微生物主要分为两类:一类是必须生活在低温条件下且最高生长温度不超过20℃、最适生长温度在15℃以下、在0℃可生长繁殖的微生物称嗜冷菌(psychrophiles);另一类其最高生长温度高于20℃、最适生长温度高于15℃、在0~5℃可生长繁殖的微生物称为耐冷菌(psychrotrophs),适合用于低温污水的生物处理。在污水生物脱氮中,耐冷菌与中温菌起着关键的作用。
1.2 低温微生物的耐低温机制
低温对细胞完整性、水黏度、溶质扩散速率、膜流动性、酶动力学以及生物细胞内外生物大分子之间的相互作用均会产生负面影响,进而对细胞功能产生不利的物理化学约束[5]。为应对低温带来的不利生长条件,低温微生物已经进化出一系列的响应机制,使其能成功地抵消伴随低温出现的各种不利生长因素,如干燥、辐射、过度紫外线、高渗透压和低营养等恶劣环境[6-7]。目前已被广泛研究的耐低温机制包括耐冷菌在低温环境中对膜流动性的保持、表达冷休克蛋白、调整酶结构、积累相容性溶质等[8-9]。此外,有研究表明微生物产生的胞外聚合物(EPS)和胞内多聚物聚羟基脂肪酸酯(PHA)也可以用来抵御寒冷[10-11]。图2是作者综合了各类文献中低温微生物应对低温生长条件所表现出的各类响应机制的研究成果[12]绘制的低温微生物假想的细胞结构。
1.2.1 胞外聚合物(EPS)

图2 综合文献研究结果所描绘的低温微生物细胞结构
生物废水处理系统中,微生物大多以聚集体的形式存在,例如污泥絮体、生物膜和颗粒污泥[13]。微生物生长繁殖过程中会分泌EPS[10],分布于细胞表面,有利于微生物细胞的凝聚。EPS的主要成分为蛋白质、多糖、核酸、磷脂及腐殖质。韩晓云[14]发现,EPS 可以在外界碳源和能量不足的情况下,为微生物的生长提供能量与碳源;此外,EPS也可以帮助微生物抵御外界环境的变化,如抗渗透压,协助微生物抵抗低温等。Ma 等[15]在研究膜污染时发现膜的污染程度随着温度的降低而逐渐加剧,这个现象从侧面反映出EPS 的含量与温度成负相关。Nichols 等[16]研究了ACM025细菌在-2℃、10℃和20℃下的EPS 产生量,发现-2℃和10℃产生的EPS量约为20℃时的30倍,ACM025具有的耐低温特性与其产生的大量EPS有一定相关性。He等[17]认为在一定温度范围内,高温或低温都将促使更多的EPS产生,因为高温能加速酶促反应,从而导致更多的EPS产生,而较低的温度则会刺激微生物分泌更多的EPS,这有利于微生物的聚集。有人在南极和北极采用宏基因组学研究发现了许多EPS生物合成基因[18]。此外,研究者还发现EPS的凝胶基质可以充当溶质的扩散屏障,通过减少溶质扩散来减小冷冻诱导的渗透压和干燥损伤[19]。迄今为止,与其他耐冷机制相比,温度对EPS影响的研究还比较少,在应对低温方面的重要作用才刚刚开始被揭开,需要进一步研究以更好地了解EPS对微生物的作用机理及其生理生态功能与组成结构的关系。
1.2.2 聚羟基脂肪酸酯
聚羟基脂肪酸酯(polyhydroxyalkanoate, PHA)是一类胞内多聚物,当碳源充足而营养元素(N、P、S、O)不足以保证细胞的正常繁殖时,细胞就会合成能量贮存颗粒[20],可以把PHA看作是一个动态能量库,当外界营养不足时,PHA 可以赋予微生物一定的生存能力和对外界各种环境变化的抵抗力。本文作者课题组研究发现,周期性地往反应器中投加高浓度的碳源能够使很多具有脱氮、除磷功能的微生物富集PHA,从而增强脱氮除磷效果[21]。Inoue 等[22]认为PHA 作为一种能量贮存物质,能够很好地抵御外界环境的变化。Ting等[23]通过蛋白质组学分析显示S. alaskensis菌株在低温条件下,与PHA 合成相关的酶的丰度会增加,以确保其在低温下保持正常的生理功能。在各种各样的PHA中,聚羟基丁酸酯(polyhydroxybutyrate, PHB)是研究最广泛的,Raiger-Iustman 等[24]分离并研究了南极菌株Pseudomonas extremaustralis,发现它能产生PHB,而PHB 正是其对抗包括寒冷在内的各种胁迫的碳源与能源功能性多聚物。目前为止,研究者已经从南极淡水和土壤中分离出了许多产PHA 的细菌菌株,这表明积累PHA 是许多细菌用来应对寒冷环境的策略之一。Sedlacek 等[25]发现细菌细胞中PHA 的存在显著增强了它们在渗透压不平衡时维持细胞完整性的能力,非嗜盐细菌Cupriavidus necator由于PHA 的存在减少了渗透压上升期间质壁分离诱导的细胞质膜损伤;在嗜盐细菌Halomonas halophila中也观察到了PHA 对低渗裂解的保护作用。以上实验结果均表明了富含PHA 的细胞能够比PHA 贫乏的细胞更有效地保持细胞完整性,PHA 能够在极端环境(低温、高渗透压和高盐度等)下保障微生物进行正常的生长代谢[26]。
2 生物脱氮工艺中功能菌的耐低温机制
温度是微生物生长最重要的因素之一,由于水温的降低,污水处理厂中微生物的生化反应速率会下降,严重影响微生物的生长代谢,进而也会影响微生物对基质的利用效率[14]。例如硝化菌在20~30℃的常温条件下,硝化速率为4.0~6.0 mgNO3ˉ-N/(gVSS·h),而在10~16℃的低温条件下,硝化速率则降至2.0~3.0 mgNO3ˉ-N/(gVSS·h)[27]。近年来,低温脱氮研究方向的相关论文数量及分类如图3 所示,其中生物膜工艺基础上衍生的各种新技术是研究的重点。相比于生物脱氮来说,低温对除磷的效果基本没有什么影响[28]。甚至有报道称,低温下的除磷效果比高温时还要好,有可能是因为低温下某些微生物的活性受到了抑制,例如聚糖菌(GAO)不能在低温下与聚磷菌(PAO)竞争碳源[29]。但也有研究表明低温对生物除磷效果有一定的影响,主要有以下两个原因,首先是在低温条件下,微生物的生长代谢速率减缓,传质速率下降;其次是低温限制了反硝化菌的活性,导致系统内硝态氮积累,使得反硝化菌与聚磷菌竞争碳源[30]。
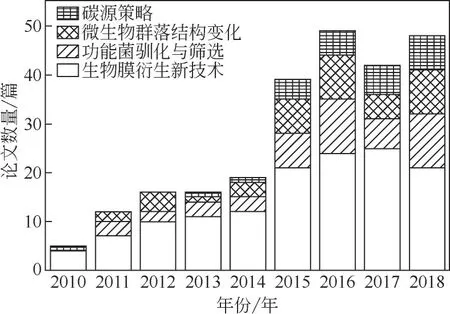
图3 2010—2018年低温生物脱氮研究方向的论文数量(Web of Science©2019 Clarivate)
2.1 AOA与AOB在废水脱氮中的作用
水中的氨态氮转化为硝酸盐的过程称为硝化反应。人们普遍认为硝化反应分为两步,氨态氮首先被氨氧化古菌(AOA)和氨氧化菌(AOB)氧化成亚硝酸盐,然后再被亚硝酸盐氧化菌(NOB)进一步氧化成硝酸盐,然而Daims 等[31]发现的全程硝化菌(comammox)推翻了100 多年来人们对硝化过程的认识,该菌直接可以将氨态氮转化为硝酸盐。有研究者对冬季污水处理厂中的AOA 与AOB 进行了检测,发现它们可以在污水处理系统中共存[32],但目前污水处理厂中comammox 的分布仍不清楚,若能清楚掌握comammox 的低温生长特性,将是低温污水生物脱氮研究的重大进展。目前已知的大多数AOA 的细胞体积普遍比AOB 的细胞小10~100倍,AOA 的氨氧化速率要比AOB 低10 倍[33]。AOA细胞特殊的四醚脂质膜比AOB 细胞膜的渗透性更低,相对于AOB,AOA 的无效离子循环量及其耗费的能量会更低,从而提供其适应极端环境的优势。此外,根据冷冻电镜中对细胞切片的断层扫描数据分析,AOA 的核糖体数量众多,使其能够对环境条件的变化(如氨浓度波动)做出快速反应。与AOB 相比,大多数AOA 能高度适应低能量的环境。因此,若能在低温或低溶解氧的废水处理系统中有效地培养和富集AOA,则可能缓解寒冷地区污水处理厂频发的硝化不良问题。
2.2 微生物群落结构的演变
温度变化对微生物群落结构和物种丰富度有着显著的影响[15]。由于温度胁迫下微生物的敏感性和抗性会有差异,低温可以改变生物废水处理系统中的微生物群落结构,甚至会降低微生物群落的多样性和物种丰富度,从而导致工艺性能恶化[34]。Zhou等[35-36]研究了三种温度(25℃、15℃、10℃)下混合A/O反应器中的微生物群落变化与脱氮效率之间的关系,结果在门水平上共观察到10 种目前已知的菌门,其中主要是变形杆菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、硝化螺旋杆菌门(Nitrospirae)和厚壁菌门(Firmicutes)。尽管它们之间的差异比较显著,但微生物群落结构在不同温度下大体还是相似的。当温度逐渐降低时,反应器中厚壁菌门的丰度一直稳定在2%以上,由于厚壁菌门在缺氧条件下可以利用硝酸盐进行反硝化,因此该混合反应器的总氮效率仍维持在一个较高的水平。Wang 等[37]分别在冬季(约7℃)和秋季(约22℃)在安徽巢湖沿岸采集沉积物样品,克隆和测序分析表明,微生物受不同环境因素的影响会呈现出不同的分布模式。大量的AOA 和厌氧氨氧化菌(anammox)表现出空间异质性,反硝化菌和异化硝酸盐还原菌(DNRA)的数量比例也会随着温度的变化而改变。以上研究结果均表明了低温对微生物结构有着显著的影响。
2.3 低温脱氮功能菌的发现
自从20世纪80年代初,Robertson和Kuenen首次从硫氧化水处理体系中分离出具有异养硝化功能的菌株以来,各国研究者相继报道了各种低温下具有脱氮功能的菌株与菌群。Zhang 等[38]在研究低温脱氮时报道过Microbacteriumsp.SFA13这种菌能在5~8℃下进行异养硝化作用。秦必达[39]在此基础上筛选出了假单胞菌属(Pseudomonassp.) 中的Pseudomonas mohnii. M-8,其在2℃下仍具有良好的脱氮性能。Huang 等[40]在实验室中分离出了Acinetobactersp. Y16,该菌株在低C/N 比和低温条件下能有效去除氨氮,但没有考察其在实际水处理工程中的稳定性。Yao 等[41]通过逐步增加溶解氧浓度成功富集和驯化了一种具有异养硝化-好氧反硝化功能的混合菌群,在10℃条件下,该菌群对氨氮和硝态氮的去除能力很高,在其富集过程中混合菌群的生物多样性和群落结构也发生了改变。系统发育分析表明,Pseudomonaspp. 和Rhodoferax ferrireducens逐渐成为富集菌群中的优势物种,其中前者在硝化-反硝化过程起主要作用,而后者则利用硝态氮作为其电子受体和氮源。路俊玲等[42]通过微生物连续富集分离,筛选得到了1株低温下具有反硝化功能的菌株,将其命名为N3,并研究了其在低温条件下的脱氮性能。结果表明,N3在C/N为8、4℃条件下,36h内即可将15mg/L硝酸盐完全去除;反硝化基因narG 和nirS 的表达与30℃下的表达量处于同一个数量级;此外,固定化N3 菌株在连续运行54 天后仍保持良好的稳定性和脱氮效果。菌株N3 在低温下具有的高效反硝化脱氮能力表明其在冬季硝酸盐水体脱氮方面具有良好的应用潜力。
3 加强低温脱氮效果的策略
调整工艺参数是保证低温脱氮工艺运行效果的首选策略。当温度降低时,通过增加水力停留时间(HRT)、污泥龄(SRT)和溶解氧(DO)等方法可以适当减轻低温带来的不利影响。此外,生物强化技术也是提高低温污水处理效果的有效方法,将特定的耐冷菌株投入到污水处理系统中,可以显著提高处理效果。近年来,生物强化技术已普遍应用于生活污水处理,来满足寒冷地区对出水排放的要求。需要注意的是,有些功能菌株需采取包埋固定措施,以确保其保持正常的生物活性[43]。有报道称,当AOB 和NOB 附着在载体生长时的活性要比悬浮生长培养的高[44],因此生物膜技术通常被用来提高低温污水处理效果,其中最常用的技术为移动床生物膜反应器(MBBR) 和曝气生物滤池(BAF)。为了进一步提高低温污水的处理效率,研究者们一直在寻找新的污水处理工艺/技术,如使用新型填料、改变碳源投加方式、投加特定的重金属等。
3.1 新型填料富集脱氮功能菌
由于生物膜法在低温时的脱氮能力较高,可根据特定的条件来选择不同的填料促进工艺的耐低温特性。生物炭具有微生物附着比表面积大、生物附着能力强和生物降解协同作用的优点。这些特点有利于提高微生物对底物的降解性能,缓解水力剪切作用,有助于维持高浓度的微生物[45]。微生物以生物炭作为载体,不仅提高了低温下的脱氮效率,还增强了复杂可溶性有机物(多糖、蛋白质、核酸等)的水解和氨化作用,促进了溶解性有机物的去除[46]。艾胜书等[47]分别通过对海绵、鲍尔环、内置海绵与鲍尔环的悬浮球生物填料序批式生物膜反应器(SBBR)处理低温污水进行了实验研究。结果表明,在10℃条件下,内置海绵与鲍尔环的悬浮球生物填料处理效果最佳,COD、-N、TN、TP的平均去除率分别为87%、69%、60%、53%,且系统稳定,启动速度快。但上述方法目前还处于实验室小试阶段,能否投入实际应用还有待证实。
3.2 投加碳源改善功能菌耐低温特性
相容性溶质(CSs)是一种低分子量的有机化合物,其在细胞内发挥作用不需要酶进行特殊适应[48]。CSs 不仅可以调节高渗透压,还可以缓解低温、冷冻等环境压力[49]。甘氨酸是CSs 的其中一种,Li等[50]通过在SBR中投加甘氨酸的研究,发现在15℃下投加了0.8mmol/L 甘氨酸的Candidatus Brocadia sinica具有良好的脱氮能力。苑宏英等[51]研究了不同碳源(丙酸钠、甲醇、乙醇及乙酸钠)对低温投加氧化还原介体1,2-萘醌-4-磺酸盐(NQS)污水生物反硝化脱氮过程的影响,发现把丙酸钠作为碳源时的反硝化速率最高,达到了7mgNOxˉ-N/(gVSS·h),分别是甲醇、乙醇和乙酸钠为碳源时的8 倍、2.6 倍和3.6 倍;硝态氮的最大去除率为61.5%,分别是甲醇、乙醇和乙酸钠为碳源时的6.9倍、9.3倍和4倍;总氮去除率为47.4%,分别是甲醇、乙醇和乙酸钠为碳源时的5.2 倍、4.6 倍和4.6倍。以上研究均表明适合的碳源投加方式有助于解决低温污水脱氮效率低的问题。
3.3 特定重金属增强功能菌活性
铜(Ⅱ)、锌(Ⅱ)和铁(Ⅱ)是富氮废水中最常见的金属元素[52],也是微生物生存所必需的微量营养素[53]。Zhang 等[54]研究了以上三种重金属对厌氧氨氧化细菌(AAOB)脱氮、生物活性及群落结构的影响,结果发现在15℃左右往氨氮和亚硝态氮浓度都为50mg/L的进水中投加1mg/L重金属后,短期暴露在1mg/L 的铜(Ⅱ)和锌(Ⅱ)中对AAOB 是有益的,这是由于以上三种金属元素是微生物生长活动所必需的;而长期暴露在1mg/L的铜(Ⅱ)和锌(Ⅱ)中会使AAOB的活性受到抑制,降低其脱氮效率,这是由于EPS虽然能够吸附金属离子,阻碍金属离子进入细胞内部[55-56],但当EPS 承受的金属离子达到饱和时,金属离子会进入细胞,破坏蛋白质结构并引起酶的代谢紊乱。铁(Ⅱ)在短期实验中对AAOB 有轻微的抑制作用,但在长期接触后会显著增强氮的去除效果,因为EPS 对铁(Ⅱ)的吸附能力高于铜(Ⅱ)和锌(Ⅱ),因此不像铜(Ⅱ)和锌(Ⅱ)那样会限制AAOB 的活性。铜(Ⅱ)和锌(Ⅱ)使具有脱氮功能的Candidatus Kuenenia的丰度从7.99%降低至3.32%和3.80%,而铁(Ⅱ)则将其丰度提高至11.30%。如何通过投加一定量的重金属来提高低温下的污水处理效果还处于起步阶段,有待更多实验数据的支持。
4 结语与展望
虽然低温脱氮研究已经取得了一定的进展,但还存在以下几个问题:①目前大多数研究成果主要还是停留在实验室小试阶段,缺乏实际工程运行的数据积累;②研究的水体主要是城市生活污水这方面,很少对工业废水或垃圾渗滤液等高浓度氨氮废水的研究;③菌种的研究大多集中在单个菌株,单一菌种很难满足各种极端环境,若能将各功能菌混合在一起研究,效果应该会更好。
任何一种新工艺/技术都有其独特的优势,若能充分利用两种或两种以上的工艺/技术,这将大大提高污水处理性能,如A2/O 与曝气生物滤池(BAF)的结合已被证实能在低温下达到高效脱氮除磷的目的[57]。在低温条件下高效处理污水还有很长一段路要走。在未来的研究工作中,可以从生物选择、特异性功能菌的固定与使用、生物倍增技术与工程实际的结合来实现在较低成本下优化各种运行参数以提高低温污水的处理效果。
